Embryologie
Embryologisch entwickelt sich das vestibulocochleäre (akustovestibuläre) Ganglion zunächst verschmolzen mit dem Ganglion des VII. Die Trennung der 3 Komponenten erfolgt allmählich. Die Neuroblasten der vestibulären und cochleären (akustischen) Ganglien stammen fast ausschließlich aus dem Epithel der Otozyste, im Gegensatz zu anderen sensorischen Ganglien des Schädels, bei denen sowohl ganglionäre als auch Neuralleisten-Plakoden umfangreiche Beiträge zur Neuroblastenpopulation leisten. Die unterstützenden Schwann- und Satellitenzellen sind jedoch, wie in allen Hirnganglien, vollständig von der Neuralleiste abstammend, offenbar aus dem Ganglion des Gesichtsnervs (siehe Abbildung unten).
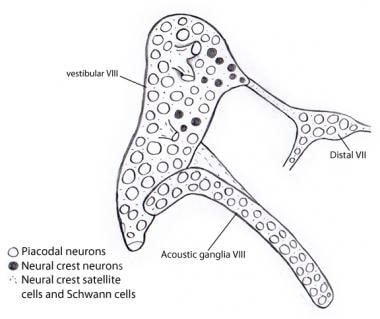 Die Abbildung zeigt, dass fast alle Neuronen in den vestibulären und akustischen Ganglien aus der Otozyste als Placodalneuronen stammen. Im Gegensatz dazu stammen alle Stütz- und Schwann-Zellen von Neuralleistenzellen ab, möglicherweise vom VII. Nervenganglion, an das das vestibulocochleäre Ganglion ursprünglich angeschlossen ist.
Die Abbildung zeigt, dass fast alle Neuronen in den vestibulären und akustischen Ganglien aus der Otozyste als Placodalneuronen stammen. Im Gegensatz dazu stammen alle Stütz- und Schwann-Zellen von Neuralleistenzellen ab, möglicherweise vom VII. Nervenganglion, an das das vestibulocochleäre Ganglion ursprünglich angeschlossen ist. Aufsteigende (afferente oder projektive) Bahnen des Hörnervs
Die aufsteigende Bahn überträgt Impulse vom Spiralorgan (von Corti) zur Großhirnrinde (siehe folgende Abbildung).
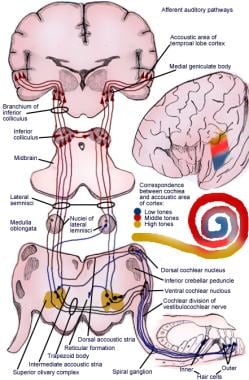 Illustration der afferenten Hörreflexbahn.
Illustration der afferenten Hörreflexbahn. Neuronen erster Ordnung des auditorischen Systems
Die Fasern des Nervus cochlearis entspringen einer Ansammlung von Nervenzellkörpern im Spiralganglion, das sich im Modiolus der Cochlea befindet. Die Neuronen des Spiralganglions sind die ersten von 4 Ordnungsneuronen zwischen der Cochlea und dem Großhirn. Sie sind bipolare Zellen, weil sie zwei Sätze von Fortsätzen oder Fasern haben, die von entgegengesetzten Enden der Zellkörper ausgehen. Die längeren zentralen Fasern, auch primäre Hörfasern genannt, bilden den Nervus cochlearis, während die kürzeren, peripheren Fasern zu den Basen der inneren und äußeren Haarzellen verlaufen. Sie verlaufen radial vom Spiralganglion bis zur Habenula perforata, einer Reihe winziger Löcher unterhalb der inneren Haarzellen. An diesem Punkt werden sie demyelinisiert und treten in das Spiralorgan (von Corti) ein.
Es gibt nur etwa 30.000 dieser Fasern, und die meisten von ihnen – etwa 95 % – versorgen die inneren Haarzellen. Der Rest durchquert den Corti-Tunnel und innerviert die äußeren Haarzellen. Die längeren zentralen Fortsätze der bipolaren Cochlea-Neuronen vereinigen sich zum Cochlea-Nervenstamm. Diese primären Hörfasern verlassen den Modiolus durch den Meatus internus und treten in die Medulla oblongata ein. Die „Mündung“ des inneren Gehörgangs (IAC) wird Porus acusticus genannt. Die anatomische Beziehung des VII. und VIII. Nervs im IAC und im Bereich des Kleinhirnbrückenwinkels sind wichtige anatomische Bereiche für die Schädelbasischirurgie und für Neurootologen.
Der VII. und VIII. Nerv sind während ihres gesamten intrakraniellen Verlaufs von Gliagewebe umschlossen. Diese Nerven sind von Schwann-Zellen umgeben, die in der IAC in der Nähe des Porus acusticus beginnen. Die Obersteiner-Redlich-Zone ist die Glia-Schwann-Grenze.
Das Scarpa-Ganglion (Vestibularganglion) liegt ungefähr in der Mitte der IAC. Die Aufteilung des Nervus cranialis (CN) VIII in den cochleären und vestibulären Ast kann im medialen Segment der IAC oder im Subarachnoidalraum erfolgen. Die hintere Hälfte der IAC wird von den superioren und inferioren Ästen des Nervus vestibularis eingenommen. Der Nervus cochlearis befindet sich anteroinferior im Kanal. Der Nervus VII. befindet sich im anterosuperioren Teil der IAC. Ein vertikaler Kamm (Bill bar) trennt den Nervus facialis und den Nervus vestibularis superior im oberen Teil der IAC. Der Querkamm trennt diesen oberen Teil der IAC von dem unteren Teil, der die unteren vestibulären und cochleären Abteilungen enthält.
Der Kleinhirnbrückenwinkel ist ein potentieller Raum in der hinteren Schädelgrube. Seine Grenzen sind wie folgt:
-
Anterior: Hintere Fossa des Schläfenbeins
-
Posterior: Vordere Oberfläche des Kleinhirns
-
Medial: Inferiore Olive
-
Oben: Unterer Rand der Pons und des Kleinhirnstiels
-
Unten: Die Kleinhirntonsille
Der Nervus trigeminus ist oberhalb des Kleinhirnwinkels sichtbar, während der IX., X. und XI. Nerv nach unten verlaufen. Weitere wichtige Strukturen im Kleinhirnbrückenwinkel sind die Arteria anterior inferior cerebellaris (AICA), der Flocculus und die seitliche Öffnung des vierten Ventrikels (Foramen Luschka). Die Arteria labyrinthina ist in der Regel ein Ast der AICA und versorgt die Cochlea und das Labyrinth.
Nach dem Eintritt in das Rückenmark ziehen die Nervenfasern der Cochlea zum Nucleus cochlearis. Der Nucleus cochlearis besteht aus fünf verschiedenen Zelltypen, die jeweils unterschiedliche morphologische und physiologische Merkmale aufweisen, wie z. B. die Reaktion auf den Beginn eines Reizes, den Offset eines Reizes und die Frequenzmodulation. Der Nucleus cochlearis ist in einen dorsalen und einen ventralen Teil unterteilt. Die Cochlea-Fasern teilen sich in 2 Hauptbündel auf: Eine Gruppe verläuft lateral und dorsal zum Restiformkörper, die andere Gruppe bleibt leicht ventral und medial zum Restiformkörper und endet im ventralen Nucleus cochlearis. Es wurde festgestellt, dass die Fasern, die aus den basalen Windungen der Cochlea stammen, im dorsalen Teil des dorsalen Cochlea-Kerns enden. Die Fasern aus den apikalen Teilen der Cochlea enden im ventralen Teil des dorsalen Cochlear Nucleus und im ventralen Nucleus. Einige Fasern gehen jedoch zu Neuronen höherer Ordnung weiter, bevor sie synapsen.
Neuronen zweiter, dritter und vierter Ordnung des auditorischen Systems
Die Zellkörper der Neuronen zweiter Ordnung liegen in den dorsalen und ventralen Kernen der Cochlea. Einige Fasern aus dem ventralen Cochlea-Kern verlaufen über die Mittellinie zu den Zellen des oberen Olivenkomplexes, während andere eine Verbindung zu den Olivenzellen derselben Seite herstellen. Der obere Olivenkomplex gilt als das erste Zentrum im aufsteigenden Hörsystem, in dem die Eingänge aus beiden Ohren zusammenlaufen. Zusammen bilden diese Fasern den trapezförmigen Körper (wo sich die Neuronen dritter Ordnung befinden).
Fasern aus dem dorsalen Cochlea-Kern überqueren die Mittellinie und enden an den Zellen der Kerne des lateralen Lemniskus. Dort treffen sie auf die Fasern aus den ventralen Cochlea-Kernen beider Seiten und aus dem Olivary-Komplex. Die auditorischen Kerne oberhalb des superioren Olivenkomplexes können exzitatorisch oder inhibitorisch sein und Inputs von jedem Ohr erhalten. Der Lemniscus ist ein wichtiger Trakt, dessen Fasern größtenteils im Colliculus inferior, dem Hörzentrum des Mittelhirns, enden.
Der Colliculus inferior ist ein komplexer Kern mit mindestens 18 Hauptzelltypen und mindestens 5 Spezialgebieten. Er ist an allen Bereichen des Hörverhaltens beteiligt, einschließlich der unterschiedlichen Empfindlichkeit für Frequenz und Intensität, Lautstärke und binaurales Hören. Obwohl einige Fasern den Colliculus umgehen und zusammen mit den Fasern aus dem Colliculus auf der nächsthöheren Ebene enden können, befinden sich die Neuronen vierter Ordnung im medialen Geniculus des Thalamus. Vom medialen genikulären Körper gibt es eine geordnete Projektion von Fasern zu einem Teil des Kortex des Schläfenlappens.
Höreingabe
Der primäre auditorische Kortex ist die erste Region der Großhirnrinde, die Höreingaben erhält. Beim Menschen und anderen Primaten ist der primäre akustische Bereich in der Großhirnrinde die obere transversale Gyri temporalis von Heschl, ein Grat im Schläfenlappen, an der Unterlippe der tiefen Spalte zwischen dem Schläfen- und dem Scheitellappen, die als laterale Sulcus (Sylvianspalte) bekannt ist.
Da etwa die Hälfte der Fasern der Hörbahnen die Mittellinie überqueren, während andere auf der gleichen Seite des Gehirns aufsteigen, ist jedes Ohr sowohl im rechten als auch im linken Kortex vertreten. Aus diesem Grund kann das binaurale Hören minimal beeinträchtigt sein, selbst wenn der auditorische Kortikalbereich einer Seite durch ein Trauma oder einen Schlaganfall verletzt wird.
Die Wahrnehmung von Schall ist mit dem rechten posterioren Gyrus temporalis superior (STG) verbunden. Der Gyrus temporalis superior enthält mehrere wichtige Strukturen des Gehirns, darunter Brodmann 41 und 42, die den Sitz des primären auditorischen Kortex markieren, der kortikalen Region, die für die Wahrnehmung grundlegender Merkmale von Geräuschen wie Tonhöhe und Rhythmus verantwortlich ist.
Das auditorische Assoziationsgebiet befindet sich im Schläfenlappen des Gehirns, in einem Gebiet, das als Wernicke-Areal oder Gebiet 22 bezeichnet wird. Dieses Gebiet in der Nähe des lateralen zerebralen Sulcus ist eine wichtige Region für die Verarbeitung von akustischen Signalen, so dass sie als Sprache, Musik oder Geräusch unterschieden werden können.
Wie bei thalamokortikalen Verbindungen üblich, empfangen Kerne im medialen genikulären Körper, die Fasern zum auditorischen Kortex senden, auch Fasern aus demselben Bereich des Kortex. Hörstörungen aufgrund bilateraler kortikaler Verletzungen, die beide Hörbereiche betreffen, wurden zwar berichtet, sind aber extrem selten. Es hat sich jedoch gezeigt, dass bilaterale Läsionen des Schläfenlappens weitreichende Auswirkungen haben (kortikale Taubheit, bei der mehrere Verhaltensweisen beeinträchtigt sind, einschließlich Sprachunterscheidung, Lokalisierung von Geräuschen und Erkennung von schwachen, kurz andauernden Signalen).
Die folgende Abbildung zeigt die bewusste und die reflexive afferente Hörbahn.
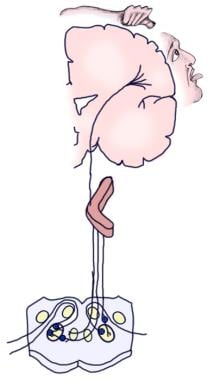 Illustration der bewussten Hörbahn.
Illustration der bewussten Hörbahn. 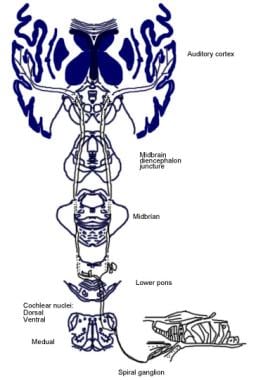 Darstellung der Gehörreflexbahn.
Darstellung der Gehörreflexbahn. Absteigende Bahnen des Hörnervs
Neben den bewussten und reflexiven afferenten Hörbahnen gibt es auch absteigende efferente Bahnen. Efferente Projektionen vom Gehirn zur Cochlea spielen ebenfalls eine Rolle bei der Wahrnehmung von Schall. Im Allgemeinen haben die absteigenden Bahnen eine hemmende Wirkung auf die aufsteigenden Fasern und dienen der Selbstregulierung des auditorischen Systems. Man geht davon aus, dass jede Relais-Hörstation doppelt innerviert wird, so dass eingehende Impulse intern beeinflusst, modifiziert oder negiert werden können.
Parallel zu der Bahn, die von den Cochlea-Kernen zum Kortex aufsteigt, gibt es eine Bahn, die vom Kortex zu den Cochlea-Kernen absteigt. In beiden Bahnen verbleiben einige der Fasern auf derselben Seite, während andere die Mittellinie zur gegenüberliegenden Seite des Gehirns überqueren. Es gibt auch Hinweise auf eine „Sporn“-Linie, die vom dorsalen Cochlea-Kern zum Kleinhirn aufsteigt, und eine weitere, die vom Colliculus inferior zum Kleinhirn absteigt.
Die Bedeutung dieser zerebralen Verbindungen ist nicht klar, aber sie könnten der evolutionären Entwicklung der Großhirnrinde vorausgehen. Vom superioren Olivenkomplex, einer Region in der Medulla oblongata, geht auch eine Faserbahn aus, die als Olivocochleäres Bündel bezeichnet wird (siehe Abbildung unten). Es stellt ein efferentes System oder eine Rückkopplungsschleife dar, über die Nervenimpulse, von denen man annimmt, dass sie hemmend wirken, die Haarzellen erreichen.
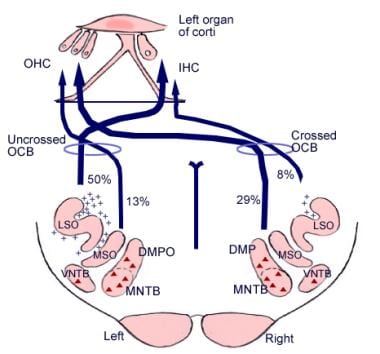 Illustration der absteigenden Bahnen des Hörnervs. Die Kreuze stellen kleine Neuronen des olivocochleären Bündels (OCB) dar; die Dreiecke stellen große Neuronen des OCB dar. DMP, DMPO = dorsaler medialer präolivärer Kern; IHC = innere Haarzelle; LSO = lateraler superiorer olivärer Kern; MNTB = medialer Kern des Trapezoidkörpers; MSO = medialer superiorer olivärer Kern; OHC = äußere Haarzelle; VNTB = ventraler Kern des Trapezoidkörpers.
Illustration der absteigenden Bahnen des Hörnervs. Die Kreuze stellen kleine Neuronen des olivocochleären Bündels (OCB) dar; die Dreiecke stellen große Neuronen des OCB dar. DMP, DMPO = dorsaler medialer präolivärer Kern; IHC = innere Haarzelle; LSO = lateraler superiorer olivärer Kern; MNTB = medialer Kern des Trapezoidkörpers; MSO = medialer superiorer olivärer Kern; OHC = äußere Haarzelle; VNTB = ventraler Kern des Trapezoidkörpers. Mikroskopische Anatomie
Siehe Innenohr für Details zur mikroskopischen Anatomie und Innervation der Zellen im Spiralorgan (von Corti) . Die folgenden Bilder zeigen diesen Bereich.
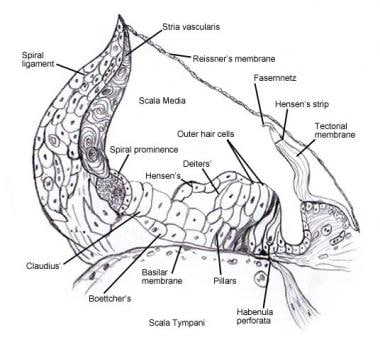 Spiralorgan (von Corti).
Spiralorgan (von Corti).  Rasterelektronenmikroskopische Aufnahme der oberen Oberfläche des Corti-Organs nach Entfernung der Tektorialmembran. Es gibt 3 Reihen äußerer Haarzellen mit ihren charakteristischen V-förmig angeordneten Stereozilien. Eine einzelne Reihe innerer Haarzellen mit einer leicht gebogenen Reihe von Stereozilien ist ebenfalls zu sehen.
Rasterelektronenmikroskopische Aufnahme der oberen Oberfläche des Corti-Organs nach Entfernung der Tektorialmembran. Es gibt 3 Reihen äußerer Haarzellen mit ihren charakteristischen V-förmig angeordneten Stereozilien. Eine einzelne Reihe innerer Haarzellen mit einer leicht gebogenen Reihe von Stereozilien ist ebenfalls zu sehen. 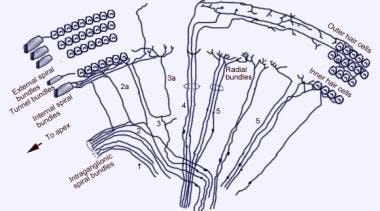 Das Muster der afferenten Innervation der Cochlea. Die Darstellung erfolgt durch die Vestibularmembran (Reissner-Membran) mit Blick „nach unten“ auf das Spiralorgan (von Corti). Die Hauptfaserbündel sind 1 und 2; 2a und 3a sind interne Spiralfasern; 4 sind externe Spiralfasern; 5 und 6 sind Radialfasern, die die inneren Haarzellen innervieren.
Das Muster der afferenten Innervation der Cochlea. Die Darstellung erfolgt durch die Vestibularmembran (Reissner-Membran) mit Blick „nach unten“ auf das Spiralorgan (von Corti). Die Hauptfaserbündel sind 1 und 2; 2a und 3a sind interne Spiralfasern; 4 sind externe Spiralfasern; 5 und 6 sind Radialfasern, die die inneren Haarzellen innervieren.