Embryology
Embryologically, the vestibulocochlear (acousticovestibular) ganglion initially develops fused with the VIIth nerve (facial) ganglion. Oddzielenie tych 3 elementów następuje stopniowo. Neuroblasty zwojów przedsionkowych i ślimakowych (akustycznych) wywodzą się prawie wyłącznie z nabłonka otocystów, w przeciwieństwie do innych zwojów czuciowych czaszki, w których zarówno zwoje, jak i grzebienie nerwowe wnoszą duży wkład w populacje neuroblastów. Jednakże wspierające komórki Schwanna i komórki satelitarne, tak jak we wszystkich zwojach czaszkowych, są w całości pochodzenia grzebienia nerwowego, najwyraźniej powstając ze zwoju nerwu twarzowego (patrz obrazek poniżej).
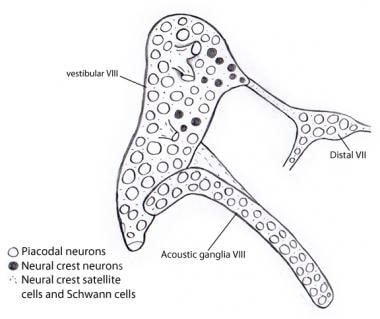 Ilustracja pokazująca, że prawie wszystkie neurony w zwojach przedsionkowych i akustycznych pochodzą z otocysty jako neurony placodalne. W przeciwieństwie do nich, wszystkie komórki podporowe i komórki Schwanna wywodzą się z komórek grzebienia nerwowego, prawdopodobnie z VII zwoju nerwowego, do którego początkowo przyczepia się zwoje przedsionkowo-ślimakowe.
Ilustracja pokazująca, że prawie wszystkie neurony w zwojach przedsionkowych i akustycznych pochodzą z otocysty jako neurony placodalne. W przeciwieństwie do nich, wszystkie komórki podporowe i komórki Schwanna wywodzą się z komórek grzebienia nerwowego, prawdopodobnie z VII zwoju nerwowego, do którego początkowo przyczepia się zwoje przedsionkowo-ślimakowe. Drogi wstępujące (aferentne lub projekcyjne) nerwu słuchowego
Droga wstępująca przekazuje impulsy z narządu spiralnego (of Corti) do kory mózgowej (patrz poniższy rysunek).
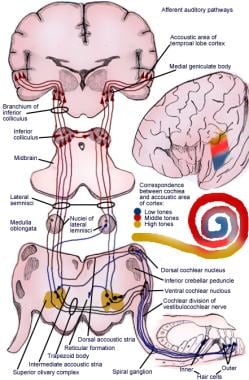 Ilustracja drogi aferentnej odruchu słuchowego.
Ilustracja drogi aferentnej odruchu słuchowego. Neurony pierwszego rzędu układu słuchowego
Włókna nerwu ślimakowego wywodzą się z agregacji ciał komórek nerwowych w zwoju spiralnym, znajdującym się w modiolusie ślimaka. Neurony zwoju spiralnego są pierwszymi neuronami 4 rzędu pomiędzy ślimakiem a mózgiem. Są to komórki dwubiegunowe, ponieważ posiadają 2 zestawy wyrostków lub włókien, które wychodzą z przeciwległych końców ciał komórkowych. Dłuższe włókna centralne, zwane również pierwotnymi włóknami słuchowymi, tworzą nerw ślimakowy, a krótsze, obwodowe włókna rozciągają się do podstaw komórek włosowatych wewnętrznych i zewnętrznych. Rozciągają się one promieniście od zwoju spiralnego do habenuli perforata, serii maleńkich otworów znajdujących się pod wewnętrznymi komórkami włosowymi. W tym miejscu, stają się zdemielinizowane, aby wejść do narządu spiralnego (of Corti).
Tylko około 30 000 tych włókien istnieje, a większa ich liczba – około 95% – unerwia wewnętrzne komórki włoskowate. Pozostałe przekraczają tunel Cortiego, aby unerwić zewnętrzne komórki włoskowate. Dłuższe wyrostki centralne dwubiegunowych neuronów ślimakowych łączą się, tworząc pień nerwowy ślimaka. Te pierwotne włókna słuchowe opuszczają modiolus przez otwór słuchowy wewnętrzny i wchodzą do rdzenia przedłużonego. Ujście” przewodu słuchowego wewnętrznego (IAC) jest nazywane porus acusticus. Anatomiczne relacje VII i VIII nerwów w IAC i regionie kąta móżdżkowo-przyśrodkowego są ważnymi obszarami anatomicznymi związanymi z chirurgią podstawy czaszki i neuro-otologami.
Nerwy VII i VIII są otoczone tkanką glejową w całym ich wewnątrzczaszkowym przebiegu. Te nerwy są otoczone przez komórki Schwanna zaczynające się w IAC blisko porus acusticus. Strefa Obersteinera-Redlicha jest połączeniem glejowo-Schwanna.
Zwoje Scarpa (przedsionkowe) leżą w przybliżeniu w środku IAC. Podział nerwu czaszkowego (CN) VIII na gałąź ślimakową i przedsionkową może nastąpić w przyśrodkowym segmencie IAC lub w przestrzeni podpajęczynówkowej. Tylna połowa IAC jest zajęta przez gałęzie górną i dolną nerwu przedsionkowego. Nerw ślimakowy znajduje się przednio-dolnie w kanale. Nerw VII znajduje się w przednio-górnej części IAC. Pionowy grzebień (Bill bar) oddziela nerw twarzowy i przedsionkowy górny w górnej części IAC. Grzebień poprzeczny oddziela tę górną część IAC od dolnej części zawierającej dolne podziały przedsionkowe i ślimakowe.
Kąt móżdżkowo-przyśrodkowy jest potencjalną przestrzenią w tylnym dole czaszki. Jego granice są następujące:
-
Zewnątrz: Tylny dół kości skroniowej
-
Przednio: Przednia powierzchnia móżdżku
-
Przyśrodkowo: Oliwka dolna
-
Przyśrodkowo: Dolna granica móżdżku i szypuły móżdżku
-
Przyśrodkowo: Migdałek móżdżku
Nerw trójdzielny jest widoczny powyżej kąta móżdżkowo-przyśrodkowego, podczas gdy nerwy IX, X i XI przebiegają w kierunku dolnym. Inne ważne struktury w obrębie kąta móżdżkowo-przyśrodkowego to tętnica móżdżkowa przednia dolna (AICA), flocculus i otwór boczny komory czwartej (foramen of Luschka). Tętnica labiryntowa jest zwykle odgałęzieniem AICA i zaopatruje ślimak i labirynt.
Po wejściu do rdzenia, włókna nerwowe ślimaka przechodzą do jądra ślimakowego. Jądro ślimakowe składa się z 5 odrębnych typów komórek, z których każdy posiada odrębne cechy morfologiczne i fizjologiczne, takie jak odpowiedź na początek bodźca, przesunięcie bodźca i modulację częstotliwości. Jądro ślimaka jest podzielone na część grzbietową i brzuszną. Włókna ślimakowe dzielą się na 2 główne wiązki: Jedna grupa przechodzi bocznie i grzbietowo do ciała spoczynkowego; druga grupa pozostaje nieco brzusznie i przyśrodkowo w stosunku do ciała spoczynkowego i kończy się w brzusznym jądrze ślimaka. Stwierdzono, że włókna pochodzące z zwojów podstawnych ślimaka kończą się w części grzbietowej jądra ślimakowego grzbietowego. Włókna z części koniuszkowych ślimaka kończą się w części brzusznej jądra ślimakowego grzbietowego i jądra brzusznego. Jednakże, niektóre włókna przechodzą do neuronów wyższego rzędu dalej wzdłuż drogi zanim synapse.
Neurony drugiego, trzeciego i czwartego rzędu układu słuchowego
Ciała komórkowe neuronów drugiego rzędu leżą w grzbietowej i brzusznej części jąder ślimaka. Niektóre włókna z brzusznego jądra ślimakowego przechodzą przez linię pośrodkową do komórek kompleksu oliwkowego górnego, podczas gdy inne łączą się z komórkami oliwkowymi tej samej strony. Kompleks oliwkowy górny jest uważany za pierwszy ośrodek wstępującego układu słuchowego, w którym zbiegają się wejścia z obu uszu. Razem włókna te tworzą ciało trapezowate (gdzie znajdują się neurony trzeciego rzędu).
Włókna z grzbietowego jądra ślimaka przecinają linię pośrodkową i kończą się na komórkach jąder lemnisk bocznych. Tam łączą się z włóknami z obustronnych jąder brzusznych ślimaka oraz z kompleksu oliwkowego. Jądra słuchowe nad kompleksem oliwkowym górnym mogą być pobudzające lub hamujące z wejściami z każdego ucha. Lemniscus jest głównym traktem, którego większość włókien kończy się w kolliculi dolnej, centrum słuchowym śródmózgowia.
Kollicula dolna jest złożonym jądrem z co najmniej 18 głównymi typami komórek i co najmniej 5 obszarami specjalizacji. Jest on zaangażowany we wszystkich obszarach zachowania słuchowego, w tym zróżnicowanej wrażliwości na częstotliwość i intensywność, głośność i obuuszne słyszenie. Chociaż niektóre włókna mogą omijać kolliculus i kończyć się, wraz z włóknami z kolliculusa, na kolejnym wyższym poziomie, przyśrodkowe ciało modzelowate wzgórza jest miejscem, gdzie znajdują się neurony czwartego rzędu. Od przyśrodkowego ciała modzelowatego, istnieje uporządkowana projekcja włókien do części kory płata skroniowego.
Wejście słuchowe
Pierwotna kora słuchowa jest pierwszym regionem kory mózgowej, który otrzymuje wejście słuchowe. U ludzi i innych naczelnych, podstawowym obszarem akustycznym w korze mózgowej jest górna poprzeczna bruzda skroniowa Heschla, grzbiet w płacie skroniowym, na dolnej wardze głębokiej szczeliny między płatami skroniowymi i ciemieniowymi, znanej jako bruzda boczna (szczelina Sylwana).
Ponieważ około połowa włókien dróg słuchowych przekracza linię środkową, podczas gdy inne wznoszą się po tej samej stronie mózgu, każde ucho jest reprezentowane zarówno w prawej, jak i lewej korze. Z tego powodu, nawet gdy słuchowy obszar korowy jednej strony jest uszkodzony w wyniku urazu lub udaru, słyszenie obuuszne może być minimalnie zaburzone.
Percepcja dźwięku jest związana z prawym tylnym górnym zakrętem skroniowym (STG). Górny zakręt skroniowy zawiera kilka ważnych struktur mózgu, w tym Brodmanna 41 i 42, wyznaczające lokalizację pierwotnej kory słuchowej, regionu korowego odpowiedzialnego za odczuwanie podstawowych cech dźwięku, takich jak wysokość i rytm.
Słuchowy obszar kojarzeniowy znajduje się w płacie skroniowym mózgu, w obszarze zwanym obszarem Wernickego lub obszarem 22. Obszar ten, w pobliżu bruzdy bocznej mózgu, jest ważnym regionem dla przetwarzania sygnałów akustycznych, aby można je było odróżnić jako mowę, muzykę lub hałas.
Jak to często bywa w przypadku połączeń wzgórzowo-korowych, jądra w obrębie przyśrodkowego ciała modzelowatego, które wysyłają włókna do kory słuchowej, również otrzymują włókna z tego samego obszaru kory. Zaburzenia słuchu spowodowane obustronnym uszkodzeniem korowym obejmującym oba obszary słuchowe były opisywane, ale są niezwykle rzadkie. Wykazano jednak, że obustronne uszkodzenie lube skroniowej może powodować rozległe skutki (głuchota korowa, w której zaburzone jest kilka zachowań, w tym rozróżnianie mowy, lokalizacja dźwięku i wykrywanie słabych sygnałów o krótkim czasie trwania).
Następujące obrazy przedstawiają świadome i odruchowe aferentne drogi słuchowe.
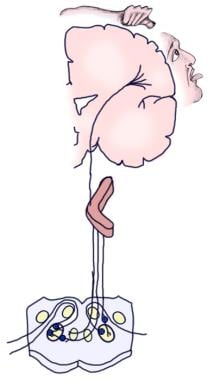 Ilustracja świadomej drogi słuchowej.
Ilustracja świadomej drogi słuchowej. 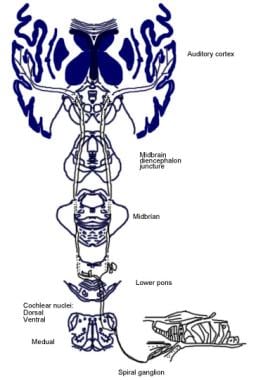 Ilustracja odruchowej drogi słuchowej.
Ilustracja odruchowej drogi słuchowej. Drogi zstępujące nerwu słuchowego
Poza świadomymi i odruchowymi drogami słuchowymi istnieją również zstępujące drogi eferentne. Projekcje eferentne z mózgu do ślimaka również odgrywają rolę w percepcji dźwięku. Ogólnie rzecz biorąc, drogi zstępujące mają hamujący wpływ na włókna wstępujące i mają tendencję do zapewnienia samoregulacji systemu słuchowego. Każda przekaźnikowa stacja słuchowa jest uważana za podwójnie unerwioną, co zapewnia drogę dla przychodzących impulsów do wewnętrznego wpływu, modyfikacji lub negacji.
Równolegle z drogą wstępującą z jąder ślimaka do kory mózgowej jest droga zstępująca z kory mózgowej do jąder ślimaka. W obu ścieżkach, niektóre włókna pozostają po tej samej stronie, podczas gdy inne przekraczają linię środkową na przeciwległą stronę mózgu. Dowody „ostrogi” linii wznoszącej się z grzbietowego jądra ślimaka do móżdżku i innego zstępującego z inferior colliculus do móżdżku również istnieją.
Znaczenie tych połączeń mózgowych nie jest jasne, ale mogą one wyprzedzać ewolucyjny rozwój kory mózgowej. Z kompleksu oliwkowego górnego, regionu w medulla oblongata, wywodzi się również trakt włóknisty zwany pęczkiem oliwkowo-ślimakowym (patrz obrazek poniżej). Stanowi on system eferentny lub pętlę sprzężenia zwrotnego, przez którą impulsy nerwowe, uważane za hamujące, docierają do komórek rzęsatych.
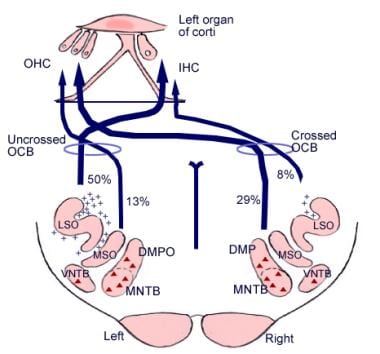 Ilustracja dróg zstępujących nerwu słuchowego. Krzyżyki reprezentują małe neurony pęczka olivocochlearnego (OCB); trójkąty reprezentują duże neurony OCB. DMP, DMPO = grzbietowe przyśrodkowe jądro przedwzrokowe; IHC = wewnętrzna komórka włosowa; LSO = boczne górne jądro oliwkowe; MNTB = przyśrodkowe jądro ciała trapezowatego; MSO = przyśrodkowe górne jądro oliwkowe; OHC = zewnętrzna komórka włosowa; VNTB = brzuszne jądro ciała trapezowatego.
Ilustracja dróg zstępujących nerwu słuchowego. Krzyżyki reprezentują małe neurony pęczka olivocochlearnego (OCB); trójkąty reprezentują duże neurony OCB. DMP, DMPO = grzbietowe przyśrodkowe jądro przedwzrokowe; IHC = wewnętrzna komórka włosowa; LSO = boczne górne jądro oliwkowe; MNTB = przyśrodkowe jądro ciała trapezowatego; MSO = przyśrodkowe górne jądro oliwkowe; OHC = zewnętrzna komórka włosowa; VNTB = brzuszne jądro ciała trapezowatego. Anatomia mikroskopowa
See Inner Ear for details regarding the microscopic anatomy and innervation of the cells in the spiral organ (of Corti) . Poniższe obrazy przedstawiają ten obszar.
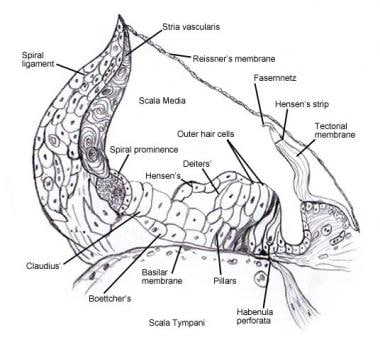 Narząd spiralny (of Corti).
Narząd spiralny (of Corti).  Skaningowy mikrograf elektronowy górnej powierzchni narządu Cortiego po usunięciu błony tektorialnej. Widoczne są 3 rzędy komórek rzęsatych zewnętrznych z charakterystycznymi stereokilami ułożonymi w kształcie litery V. Widoczny jest również pojedynczy rząd wewnętrznych komórek włoskowatych z lekko zakrzywionym rzędem stereokili.
Skaningowy mikrograf elektronowy górnej powierzchni narządu Cortiego po usunięciu błony tektorialnej. Widoczne są 3 rzędy komórek rzęsatych zewnętrznych z charakterystycznymi stereokilami ułożonymi w kształcie litery V. Widoczny jest również pojedynczy rząd wewnętrznych komórek włoskowatych z lekko zakrzywionym rzędem stereokili. 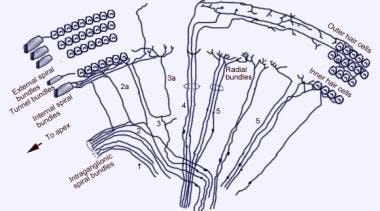 Schemat unerwienia aferentnego ślimaka. Ilustracja jest pokazana przez błonę przedsionkową (Reissnera), patrząc „w dół” na narząd spiralny (Cortiego). Główne wiązki włókien to 1 i 2; 2a i 3a to włókna spiralne wewnętrzne; 4 to włókna spiralne zewnętrzne; 5 i 6 to włókna promieniowe unerwiające wewnętrzne komórki włoskowate.
Schemat unerwienia aferentnego ślimaka. Ilustracja jest pokazana przez błonę przedsionkową (Reissnera), patrząc „w dół” na narząd spiralny (Cortiego). Główne wiązki włókien to 1 i 2; 2a i 3a to włókna spiralne wewnętrzne; 4 to włókna spiralne zewnętrzne; 5 i 6 to włókna promieniowe unerwiające wewnętrzne komórki włoskowate.