Embryologi
Embryologiskt sett utvecklas det vestibulocochleära (akustikovestibulära) gangliet till en början sammansvetsat med den VII:e nervens (ansikts) ganglion. Separationen av de 3 komponenterna sker gradvis. Neuroblasterna i de vestibulära och cochleära (akustiska) ganglierna härstammar nästan uteslutande från otocystepitelet, i motsats till andra kraniala sensoriska ganglier där både ganglionära och neuralkammarplacoder ger omfattande bidrag till neuroblastpopulationerna. De stödjande Schwann- och satellitcellerna, som i alla kraniala ganglier, har dock helt och hållet sitt ursprung i neurallistan och härstammar uppenbarligen från ansiktsnerverns ganglion (se bilden nedan).
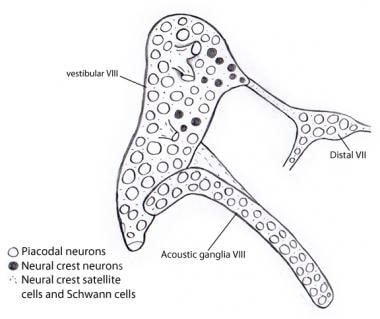 Illustration som visar att nästan alla neuroner i de vestibulära och akustiska ganglierna härstammar från otocystan som placodala neuroner. Däremot härstammar alla stödje- och Schwann-celler från neuralkamceller, möjligen från det VII:e nervganglion till vilket det vestibulocochleära ganglionet ursprungligen är knutet.
Illustration som visar att nästan alla neuroner i de vestibulära och akustiska ganglierna härstammar från otocystan som placodala neuroner. Däremot härstammar alla stödje- och Schwann-celler från neuralkamceller, möjligen från det VII:e nervganglion till vilket det vestibulocochleära ganglionet ursprungligen är knutet. Hörselnervens uppåtgående (afferenta eller projektiva) banor
Den uppåtgående banan överför impulser från spiralorganet (of Corti) till hjärnbarken (se följande bild).
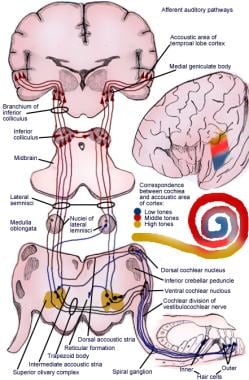 Illustration av den afferenta hörselreflexbanan.
Illustration av den afferenta hörselreflexbanan. Auditiva systemets neuroner av första ordningen
Fibrerna i cochlearisnerven har sitt ursprung i en aggregering av nervcellskroppar i spiralganglionet, som ligger i modiolus i cochlea. Neuronerna i spiralganglionet är de första av 4 ordningsneuronerna mellan cochlea och cerebrum. De är bipolära celler, eftersom de har två uppsättningar processer, eller fibrer, som sträcker sig från motsatta ändar av cellkropparna. De längre centrala fibrerna, även kallade de primära hörselfibrerna, bildar cochlearisnerven, och de kortare, perifera fibrerna sträcker sig till baserna av de inre och yttre hårcellerna. De sträcker sig radiellt från spiralganglionet till habenula perforata, en serie små hål under de inre hårcellerna. Där blir de demyeliniserade och går in i Cortis spiralorgan.
Det finns bara cirka 30 000 av dessa fibrer, och det största antalet av dem – cirka 95 % – nerverar de inre hårcellerna. Resten korsar Cortis tunnel för att nervera de yttre hårcellerna. De längre centrala processerna hos de bipolära cochleära neuronerna förenas för att bilda den cochleära nervstammen. Dessa primära auditiva fibrer lämnar modiolus genom den inre gången och går in i medulla oblongata. Den inre hörselkanalens (IAC) ”mun” kallas porus acusticus. Det anatomiska förhållandet mellan de VII:e och VIII:e nerverna i IAC och cerebellopontinvinkelregionen är viktiga anatomiska områden med anknytning till skallbaskirurgi och neurootologer.
De VII:e och VIII:e nerverna är inneslutna i gliavävnad under hela sitt intrakraniella förlopp. Dessa nerver är omgivna av Schwann-celler som börjar i IAC nära porus acusticus. Obersteiner-Redlich-zonen är glial-Schwann-övergången.
Scarpa (vestibulära) ganglion ligger ungefär i mitten av IAC. Delningen av kranialnerv (CN) VIII i de cochleära och vestibulära grenarna kan ske i det mediala segmentet av IAC eller i subarachnoidalrummet. Den bakre halvan av IAC upptas av den vestibulära nervens övre och undre grenar. Nervus cochlearis ligger anteroinferior i kanalen. Den VII:e nerven är belägen i den anterosuperiora delen av IAC. En vertikal kam (Bill bar) skiljer ansiktsnerven och den övre vestibulära nerverna åt i den övre delen av IAC. Den tvärgående kammen skiljer denna övre del av IAC från den nedre delen som innehåller de nedre vestibulära och cochleära divisionerna.
Cerebellopontinvinkeln är ett potentiellt utrymme i den bakre kranialgropen. Dess gränser är följande:
-
Anteriört: Bakre fossa i temporalbenet
-
Posterior: Bakre fossa i temporalbenet
-
Förre ytan av lillhjärnan
-
Medialt: Inferior olive
-
Superiört: Inferior gränsen av pons och cerebellar peduncle
-
Inferior: Cerebellartonsillen
Trigeminusnerven är synlig över den cerebellopontina vinkeln, medan den IX:e, X:e och XI:e nerverna löper inferiört. Andra viktiga strukturer inom den cerebellopontina vinkeln är den främre nedre cerebellära artären (AICA), flocculus och den laterala öppningen av den fjärde ventrikeln (Luschkaforamen). Labyrintinarterien är vanligtvis en gren av AICA och försörjer cochlea och labyrint.
Efter att ha kommit in i märgen fortsätter de cochleära nervfibrerna till den cochleära kärnan. Den cochleära kärnan består av fem olika celltyper, var och en med olika morfologiska och fysiologiska egenskaper, t.ex. respons på stimulusstart, stimulusförskjutning och frekvensmodulering. Den cochleära kärnan är uppdelad i den dorsala och ventrala delen. De cochleära fibrerna delas upp i två huvudbuntar: Den ena gruppen passerar lateralt och dorsalt till restiformkroppen, den andra gruppen förblir något ventral och medial till restiformkroppen och slutar i den ventrala cochlearkärnan. Fibrer som kommer från de basala spolarna i cochlea har visat sig sluta i den dorsala delen av den dorsala cochlearkärnan. Fibrerna från de apikala delarna av cochlea slutar i den ventrala delen av den dorsala cochlearkärnan och i den ventrala kärnan. Vissa fibrer passerar dock till neuroner av högre ordning längre fram i banan innan de synapserar.
Neuroner av andra, tredje och fjärde ordningen i hörselsystemet
Cellkropparna hos neuronerna av andra ordningen ligger i de dorsala och ventrala cochlearkärnorna. Vissa fibrer från den ventrala cochlearkärnan passerar över mittlinjen till cellerna i det övre olivariekomplexet, medan andra gör förbindelse med olivariecellerna på samma sida. Det övre olivariekomplexet anses vara det första centret i det uppåtgående auditiva systemet, där ingångar från båda öronen konvergerar. Tillsammans bildar dessa fibrer trapetskroppen (där tredje ordningens neuroner finns).
Fibrer från den dorsala cochlearkärnan korsar mittlinjen för att sluta på cellerna i kärnorna i laterala lemniscus. Där förenas de med fibrer från de ventrala cochlearkärnorna på båda sidor och från olivariekomplexet. Hörselkärnorna ovanför det övre olivariekomplexet kan vara excitatoriska eller hämmande med ingångar från båda öronen. Lemniscus är ett viktigt tractus, vars flesta fibrer slutar i inferior colliculus, det auditiva centret i mellanhjärnan.
Inferior colliculus är en komplex kärna med minst 18 större celltyper och minst 5 specialiseringsområden. Den är involverad i alla områden av auditivt beteende, inklusive differentiell känslighet för frekvens och intensitet, ljudstyrka och binaural hörsel. Även om vissa fibrer kan gå förbi colliculus och sluta, tillsammans med fibrerna från colliculus, på nästa högre nivå, är det i thalamus medial geniculate body i thalamus som neuronerna av fjärde ordningen är lokaliserade. Från den mediala genikulära kroppen finns en ordnad projektion av fibrer till en del av hjärnbarken i tinningloben.
Auditorisk input
Den primära auditiva cortexen är den första regionen i hjärnbarken som tar emot auditiv input. Hos människor och andra primater är det primära akustiska området i hjärnbarken Heschls övre transversala temporala gyri, en ås i tinningloben, på underläppen av den djupa klyftan mellan tinning- och parietalloberna, känd som den laterala sulcus (Sylvian fissure).
Om ungefär hälften av fibrerna i hörselbanorna korsar mittlinjen medan andra stiger upp på samma sida av hjärnan, är varje öra representerat i både höger och vänster cortex. Av denna anledning kan den binaurala hörseln påverkas minimalt även när det auditiva kortikala området på ena sidan skadas av trauma eller stroke.
Perceptionen av ljud är förknippad med den högra bakre övre temporala gyrus (STG). Den övre temporala gyrus innehåller flera viktiga strukturer i hjärnan, bland annat Brodmann 41 och 42, som markerar placeringen av den primära auditiva cortexen, den kortikala region som ansvarar för känslan av ljudets grundläggande egenskaper som tonhöjd och rytm.
Det auditiva associationsområdet ligger i hjärnans temporallob, i ett område som kallas Wernicke-området eller område 22. Detta område, nära den laterala cerebrala sulcus, är ett viktigt område för bearbetning av akustiska signaler så att de kan särskiljas som tal, musik eller buller.
Som är vanligt för thalamokortikala kopplingar, tar kärnor i den mediala genikulära kroppen som sänder fibrer till den auditiva hjärnbarken också emot fibrer från samma område i hjärnbarken. Hörselnedsättning på grund av bilateral kortikal skada som omfattar båda hörselområdena har rapporterats, men det är ytterst sällsynt. Däremot har bilaterala lesioner av den temporala loben visat sig ge omfattande effekter (kortikal dövhet, där flera beteenden påverkas, bland annat taldiskriminering, lokalisering av ljud och upptäckt av svaga, kortvariga signaler).
Följande bild skildrar de medvetna och reflexmässiga afferenta hörselbanorna.
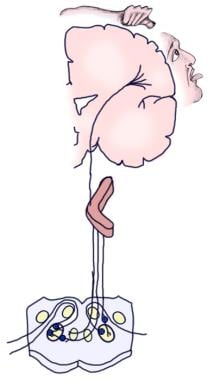 Illustration av den medvetna hörselbanan.
Illustration av den medvetna hörselbanan. 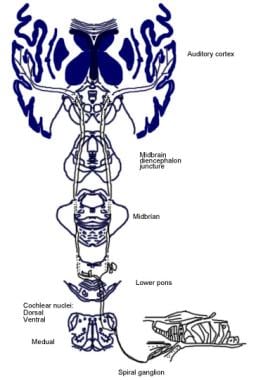 Illustration av den auditiva reflexbanan.
Illustration av den auditiva reflexbanan. Hörselnervens nedåtgående banor
Bortsett från de medvetna och reflexmässiga afferenta hörselbanorna finns det även nedåtgående efferenta banor. Efferenta projektioner från hjärnan till snäckan spelar också en roll för uppfattningen av ljud. I allmänhet har de nedåtgående banorna en hämmande effekt på de uppåtgående fibrerna, och de tenderar att ge självreglering åt det auditiva systemet. Varje reläauditiv station anses vara dubbelt innerverad, vilket ger ett sätt för inkommande impulser att internt påverkas, modifieras eller förnekas.
Parallellt med den stigande banan från cochlearkärnorna till hjärnbarken finns en nedåtgående banan från hjärnbarken till cochlearkärnorna. I båda banorna stannar en del av fibrerna på samma sida, medan andra korsar mittlinjen till den motsatta sidan av hjärnan. Det finns också belägg för en ”sporre”-linje som stiger upp från den dorsala cochlearkärnan till lillhjärnan och en annan som sjunker ner från inferior colliculus till lillhjärnan.
Betydelsen av dessa hjärnförbindelser är inte klarlagd, men de kan vara äldre än den evolutionära utvecklingen av hjärnbarken. Från det övre olivariekomplexet, en region i medulla oblongata, utgår också ett fiberstråk som kallas olivocochlear bundle (se bilden nedan). Den utgör ett efferent system, eller en återkopplingsslinga, genom vilken nervimpulser, som tros vara hämmande, når hårcellerna.
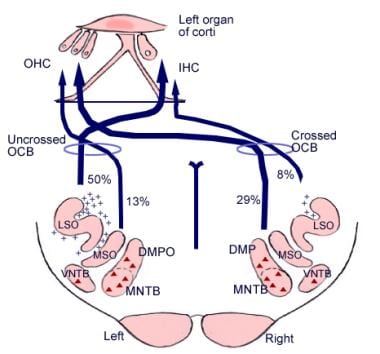 Illustration av hörselnervens nedåtgående banor. Kryssen representerar små olivocochlear bundle (OCB)-neuroner, trianglarna representerar stora OCB-neuroner. DMP, DMPO = dorsal medial preolivary nucleus; IHC = inre hårcell; LSO = lateral superior olivary nucleus; MNTB = medial nucleus of trapezoid body; MSO = medial superior olivary nucleus; OHC = yttre hårcell; VNTB = ventral nucleus of trapezoid body.
Illustration av hörselnervens nedåtgående banor. Kryssen representerar små olivocochlear bundle (OCB)-neuroner, trianglarna representerar stora OCB-neuroner. DMP, DMPO = dorsal medial preolivary nucleus; IHC = inre hårcell; LSO = lateral superior olivary nucleus; MNTB = medial nucleus of trapezoid body; MSO = medial superior olivary nucleus; OHC = yttre hårcell; VNTB = ventral nucleus of trapezoid body. Mikroskopisk anatomi
Se Innerörat för detaljer om den mikroskopiska anatomin och innervation av cellerna i Cortis spiralorgan . Följande bilder visar detta område.
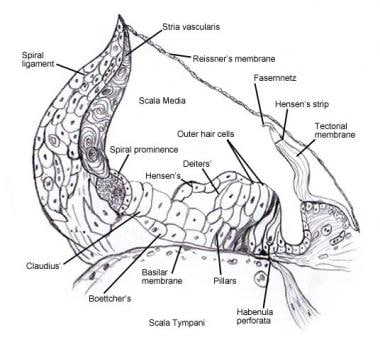 Spiralorganet (Corti).
Spiralorganet (Corti).  Skanningselektronmikroskopisk bild av den övre ytan av Corti-organet efter avlägsnande av tectorialmembranet. Det finns tre rader av yttre hårceller med sina karakteristiska V-ordnade stereocilier. En enda rad inre hårceller med en något böjd rad stereociler visas också.
Skanningselektronmikroskopisk bild av den övre ytan av Corti-organet efter avlägsnande av tectorialmembranet. Det finns tre rader av yttre hårceller med sina karakteristiska V-ordnade stereocilier. En enda rad inre hårceller med en något böjd rad stereociler visas också. 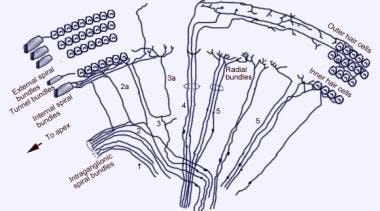 Snäckans afferenta innerveringsmönster. Illustrationen visas genom det vestibulära (Reissners) membranet och tittar ”ner” på spiralorganet (Corti). De viktigaste fiberbuntarna är 1 och 2; 2a och 3a är inre spiralfibrer; 4 är yttre spiralfibrer; 5 och 6 är radiella fibrer som innerverar de inre hårcellerna.
Snäckans afferenta innerveringsmönster. Illustrationen visas genom det vestibulära (Reissners) membranet och tittar ”ner” på spiralorganet (Corti). De viktigaste fiberbuntarna är 1 och 2; 2a och 3a är inre spiralfibrer; 4 är yttre spiralfibrer; 5 och 6 är radiella fibrer som innerverar de inre hårcellerna.