Embryologie
Embryologiquement, le ganglion vestibulocochléaire (acousticovestibulaire) se développe initialement en fusion avec le ganglion du VIIe nerf (facial). La séparation des 3 composants se fait progressivement. Les neuroblastes des ganglions vestibulaire et cochléaire (acoustique) proviennent presque exclusivement de l’épithélium de l’otocyste, contrairement aux autres ganglions sensoriels crâniens dans lesquels les placodes ganglionnaires et celles de la crête neurale contribuent largement aux populations de neuroblastes. Cependant, les cellules satellites et de Schwann de soutien, comme dans tous les ganglions crâniens, sont entièrement d’origine crête neurale, provenant apparemment du ganglion du nerf facial (voir l’image ci-dessous).
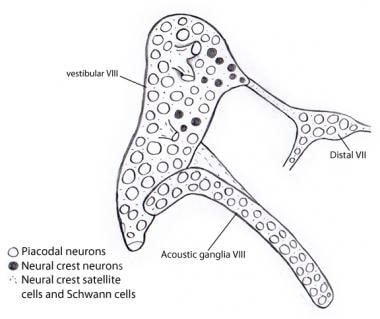 Illustration montrant que presque tous les neurones des ganglions vestibulaires et acoustiques sont dérivés de l’otocyste comme neurones placodaux. En revanche, toutes les cellules de soutien et de Schwann sont dérivées des cellules de la crête neurale, peut-être du VIIe ganglion nerveux auquel le ganglion vestibulocochléaire est initialement rattaché.
Illustration montrant que presque tous les neurones des ganglions vestibulaires et acoustiques sont dérivés de l’otocyste comme neurones placodaux. En revanche, toutes les cellules de soutien et de Schwann sont dérivées des cellules de la crête neurale, peut-être du VIIe ganglion nerveux auquel le ganglion vestibulocochléaire est initialement rattaché. Voies ascendantes (afférentes ou projectives) du nerf auditif
La voie ascendante transmet les impulsions de l’organe spiral (de Corti) au cortex cérébral (voir l’image suivante).
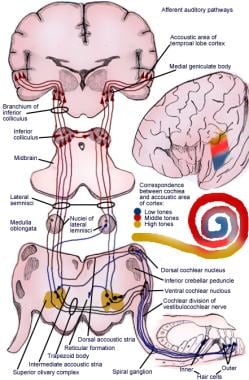 Illustration de la voie afférente du réflexe auditif.
Illustration de la voie afférente du réflexe auditif. Nerones de premier ordre du système auditif
Les fibres du nerf cochléaire proviennent d’une agrégation de corps cellulaires nerveux dans le ganglion spiral, situé dans le modiolus de la cochlée. Les neurones du ganglion spiral sont les premiers des 4 neurones d’ordre entre la cochlée et le cerveau. Ce sont des cellules bipolaires, car elles possèdent deux ensembles de processus, ou fibres, qui partent des extrémités opposées des corps cellulaires. Les fibres centrales, plus longues, également appelées fibres auditives primaires, forment le nerf cochléaire, tandis que les fibres périphériques, plus courtes, s’étendent jusqu’aux bases des cellules ciliées internes et externes. Elles s’étendent radialement depuis le ganglion spiral jusqu’à l’habenula perforata, une série de petits trous situés sous les cellules ciliées internes. A cet endroit, elles se démyélinisent pour entrer dans l’organe spiral (de Corti).
Il n’existe qu’environ 30 000 de ces fibres, et la plus grande partie d’entre elles – environ 95% – innerve les cellules ciliées internes. Les autres traversent le tunnel de Corti pour innerver les cellules ciliées externes. Les processus centraux les plus longs des neurones cochléaires bipolaires s’unissent pour former le tronc du nerf cochléaire. Ces fibres auditives primaires sortent du modiolus par le méat interne et entrent dans le bulbe rachidien. La « bouche » du conduit auditif interne (CAI) s’appelle le porus acusticus. Les relations anatomiques des VIIe et VIIIe nerfs dans le CAI et la région de l’angle ponto-cérébelleux sont des zones anatomiques importantes liées à la chirurgie de la base du crâne et aux neuro-otologues.
Les VIIe et VIIIe nerfs sont enveloppés de tissu glial tout au long de leur parcours intracrânien. Ces nerfs sont entourés de cellules de Schwann qui commencent dans le CAI près du porus acusticus. La zone d’Obersteiner-Redlich est la jonction gliale-Schwann.
Le ganglion de Scarpa (vestibulaire) se situe approximativement au milieu du CAI. La division du nerf crânien (CN) VIII en branches cochléaire et vestibulaire peut se produire dans le segment médian du CAI ou dans l’espace sous-arachnoïdien. La moitié postérieure du CAI est occupée par les branches supérieure et inférieure du nerf vestibulaire. Le nerf cochléaire est situé antéro-inférieur dans le canal. Le VIIe nerf est situé dans la partie antéro-supérieure du CAI. Une crête verticale (barre de Bill) sépare les nerfs facial et vestibulaire supérieur dans la partie supérieure du CAI. La crête transversale sépare cette partie supérieure du CAI de la partie inférieure contenant les divisions vestibulaires et cochléaires inférieures.
L’angle ponto-cérébelleux est un espace potentiel de la fosse crânienne postérieure. Ses limites sont les suivantes :
-
Antérieurement : Fosse postérieure de l’os temporal
-
Postérieurement : Surface antérieure du cervelet
-
Médialement : Olive inférieure
-
Supérieurement : Bordure inférieure du pons et du pédoncule cérébelleux
-
Inférieurement : L’amygdale cérébelleuse
Le nerf trijumeau est visible au-dessus de l’angle ponto-cérébelleux, tandis que les IXe, Xe et XIe nerfs se dirigent vers l’intérieur. D’autres structures importantes dans l’angle ponto-cérébelleux comprennent l’artère cérébelleuse inférieure antérieure (AICA), le flocculus et l’ouverture latérale du quatrième ventricule (foramen de Luschka). L’artère labyrinthique est généralement une branche de l’AICA et alimente la cochlée et le labyrinthe.
Après avoir pénétré dans la moelle, les fibres du nerf cochléaire se dirigent vers le noyau cochléaire. Le noyau cochléaire est constitué de 5 types de cellules distinctes, chacune ayant des caractéristiques morphologiques et physiologiques distinctes, comme la réponse au début du stimulus, au décalage du stimulus et à la modulation de fréquence. Le noyau cochléaire est divisé en deux parties, dorsale et ventrale. Les fibres cochléaires se divisent en 2 faisceaux principaux : Un groupe passe latéralement et dorsalement au corps réstiforme ; l’autre groupe reste légèrement ventral et médial au corps réstiforme et se termine dans le noyau cochléaire ventral. On a constaté que les fibres provenant des spires basales de la cochlée se terminent dans la partie dorsale du noyau cochléaire dorsal. Les fibres provenant des parties apicales de la cochlée se terminent dans la partie ventrale du noyau cochléaire dorsal et dans le noyau ventral. Cependant, certaines fibres passent à des neurones d’ordre supérieur plus loin dans la voie avant de faire synapse.
Nerones de deuxième, troisième et quatrième ordre du système auditif
Les corps cellulaires des neurones de deuxième ordre se trouvent dans les noyaux cochléaires dorsal et ventral. Certaines fibres du noyau cochléaire ventral traversent la ligne médiane jusqu’aux cellules du complexe olivaire supérieur, tandis que d’autres établissent une connexion avec les cellules olivaires du même côté. Le complexe olivaire supérieur est considéré comme le premier centre du système auditif ascendant, où convergent les entrées des deux oreilles. Ensemble, ces fibres forment le corps trapézoïdal (où sont situés les neurones de troisième ordre).
Les fibres du noyau cochléaire dorsal traversent la ligne médiane pour se terminer sur les cellules des noyaux du lemniscus latéral. Là, elles sont rejointes par les fibres provenant des noyaux cochléaires ventraux des deux côtés et du complexe olivaire. Les noyaux auditifs situés au-dessus du complexe olivaire supérieur peuvent être excitateurs ou inhibiteurs avec des entrées provenant de chaque oreille. Le lemniscus est un tractus majeur dont la plupart des fibres se terminent dans le colliculus inférieur, le centre auditif du mésencéphale.
Le colliculus inférieur est un noyau complexe avec au moins 18 types cellulaires majeurs et au moins 5 zones de spécialisation. Il est impliqué dans tous les domaines du comportement auditif, y compris la sensibilité différentielle pour la fréquence et l’intensité, l’intensité sonore et l’audition binaurale. Bien que certaines fibres puissent contourner le colliculus et se terminer, avec les fibres du colliculus, au niveau supérieur suivant, le corps géniculé médian du thalamus est l’endroit où se trouvent les neurones de quatrième ordre. A partir du corps géniculé médian, il y a une projection ordonnée de fibres vers une partie du cortex du lobe temporal.
Entrée auditive
Le cortex auditif primaire est la première région du cortex cérébral à recevoir une entrée auditive. Chez les humains et les autres primates, la zone acoustique primaire du cortex cérébral est la gyrie temporale transversale supérieure de Heschl, une crête du lobe temporal, sur la lèvre inférieure de la fente profonde entre les lobes temporal et pariétal, connue sous le nom de sillon latéral (fissure sylvienne).
Parce qu’environ la moitié des fibres des voies auditives traversent la ligne médiane alors que d’autres montent du même côté du cerveau, chaque oreille est représentée dans le cortex droit et gauche. Pour cette raison, même lorsque la zone corticale auditive d’un côté est lésée par un traumatisme ou un accident vasculaire cérébral, l’audition binaurale peut être peu affectée.
La perception des sons est associée au gyrus temporal supérieur postérieur droit (STG). Le gyrus temporal supérieur contient plusieurs structures importantes du cerveau, notamment Brodmann 41 et 42, marquant l’emplacement du cortex auditif primaire, la région corticale responsable de la sensation des caractéristiques de base du son telles que la hauteur et le rythme.
L’aire d’association auditive est située dans le lobe temporal du cerveau, dans une zone appelée aire de Wernicke ou aire 22. Cette zone, proche du sillon cérébral latéral, est une région importante pour le traitement des signaux acoustiques afin qu’ils puissent être distingués comme parole, musique ou bruit.
Comme cela est courant pour les connexions thalamocorticales, les noyaux du corps géniculé médian qui envoient des fibres au cortex auditif reçoivent également des fibres de la même zone du cortex. Une déficience auditive due à une lésion corticale bilatérale impliquant les deux zones auditives a été signalée, mais elle est extrêmement rare. Cependant, il a été démontré que des lésions bilatérales du lobe temporal produisent des effets de grande ampleur (surdité corticale, dans laquelle plusieurs comportements sont affectés, notamment la discrimination de la parole, la localisation du son et la détection de signaux faibles et de courte durée).
L’image suivante représente les voies auditives afférentes conscientes et réflexes.
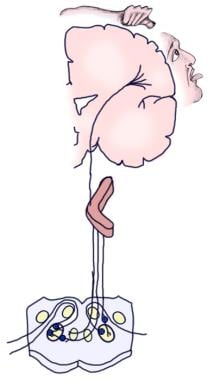 Illustration de la voie auditive consciente.
Illustration de la voie auditive consciente. 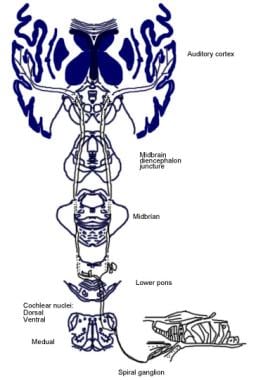 Illustration de la voie auditive réflexe.
Illustration de la voie auditive réflexe. Voies descendantes du nerf auditif
En plus des voies auditives afférentes conscientes et réflexes, il existe également des voies efférentes descendantes. Les projections efférentes du cerveau vers la cochlée jouent également un rôle dans la perception du son. En général, les voies descendantes ont un effet inhibiteur sur les fibres ascendantes, et elles tendent à assurer l’autorégulation du système auditif. Chaque station auditive relais est considérée comme doublement innervée, ce qui permet aux impulsions entrantes d’être influencées, modifiées ou annulées de façon interne.
Parallèlement à la voie ascendante des noyaux cochléaires vers le cortex, il existe une voie descendante du cortex vers les noyaux cochléaires. Dans les deux voies, certaines des fibres restent du même côté, tandis que d’autres traversent la ligne médiane vers le côté opposé du cerveau. Des preuves d’une ligne « éperon » montant du noyau cochléaire dorsal vers le cervelet et une autre descendant du colliculus inférieur vers le cervelet existent également.
La signification de ces connexions cérébrales n’est pas claire, mais elles pourraient être antérieures au développement évolutif du cortex cérébral. Du complexe olivaire supérieur, une région du bulbe rachidien, part également un tractus de fibres appelé faisceau olivocochléaire (voir l’image ci-dessous). Il constitue un système efférent, ou boucle de rétroaction, par lequel les impulsions nerveuses, que l’on pense être inhibitrices, atteignent les cellules ciliées.
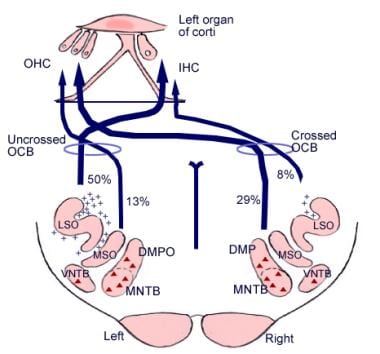 Illustration des voies descendantes du nerf auditif. Les croix représentent les petits neurones du faisceau olivocochléaire (BOC) ; les triangles représentent les gros neurones du BOC. DMP, DMPO = noyau préolivaire médial dorsal ; IHC = cellule ciliée interne ; LSO = noyau olivaire supérieur latéral ; MNTB = noyau médial du corps trapézoïdal ; MSO = noyau olivaire supérieur médial ; OHC = cellule ciliée externe ; VNTB = noyau ventral du corps trapézoïdal.
Illustration des voies descendantes du nerf auditif. Les croix représentent les petits neurones du faisceau olivocochléaire (BOC) ; les triangles représentent les gros neurones du BOC. DMP, DMPO = noyau préolivaire médial dorsal ; IHC = cellule ciliée interne ; LSO = noyau olivaire supérieur latéral ; MNTB = noyau médial du corps trapézoïdal ; MSO = noyau olivaire supérieur médial ; OHC = cellule ciliée externe ; VNTB = noyau ventral du corps trapézoïdal. Anatomie microscopique
Voir Oreille interne pour des détails concernant l’anatomie microscopique et l’innervation des cellules de l’organe spiral (de Corti) . Les images suivantes représentent cette zone.
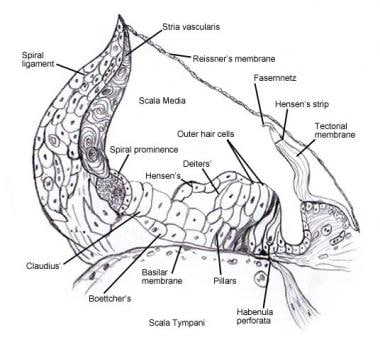 Organe spiral (de Corti).
Organe spiral (de Corti).  Micrographie électronique à balayage de la surface supérieure de l’organe de Corti après retrait de la membrane tectoriale. Il y a 3 rangées de cellules ciliées extérieures avec leurs stéréocils caractéristiques disposés en V. Une seule rangée de cellules ciliées internes avec une rangée de stéréocils légèrement courbée est également représentée.
Micrographie électronique à balayage de la surface supérieure de l’organe de Corti après retrait de la membrane tectoriale. Il y a 3 rangées de cellules ciliées extérieures avec leurs stéréocils caractéristiques disposés en V. Une seule rangée de cellules ciliées internes avec une rangée de stéréocils légèrement courbée est également représentée. 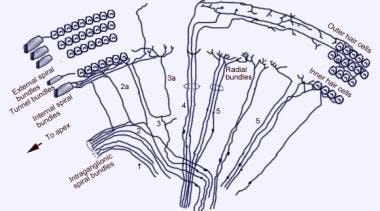 Le schéma d’innervation afférente de la cochlée. L’illustration est montrée à travers la membrane vestibulaire (de Reissner), en regardant « en bas » sur l’organe spiralé (de Corti). Les principaux faisceaux de fibres sont 1 et 2 ; 2a et 3a sont des fibres spiralées internes ; 4 sont des fibres spiralées externes ; 5 et 6 sont des fibres radiales innervant les cellules ciliées internes.
Le schéma d’innervation afférente de la cochlée. L’illustration est montrée à travers la membrane vestibulaire (de Reissner), en regardant « en bas » sur l’organe spiralé (de Corti). Les principaux faisceaux de fibres sont 1 et 2 ; 2a et 3a sont des fibres spiralées internes ; 4 sont des fibres spiralées externes ; 5 et 6 sont des fibres radiales innervant les cellules ciliées internes.