Embriologia
Embriologicamente, o gânglio vestibulococlear (acústicoovestibular) desenvolve-se inicialmente fundido com o gânglio do VII nervo (facial). A separação dos 3 componentes ocorre gradualmente. Os neuroblastos ganglionares vestibulares e cocleares (acústicos) são derivados quase exclusivamente do epitélio otocistolar, em contraste com outros gânglios sensoriais cranianos, nos quais tanto os gânglios ganglionares como os gânglios de crista neural fazem contribuições extensas para as populações de neuroblastos. Entretanto, o suporte de Schwann e células satélites, como em todos os gânglios cranianos, são inteiramente de origem neural, aparentemente surgindo a partir do gânglio do nervo facial (veja a imagem abaixo).
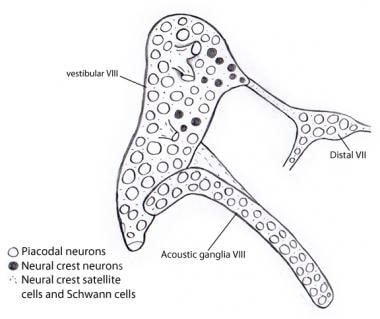 Ilustração mostrando que quase todos os neurônios nos gânglios vestibulares e acústicos são derivados do otocisto como neurônios placodais. Em contraste, todas as células de suporte e Schwann são derivadas das células da crista neural, possivelmente do VII gânglio nervoso ao qual o gânglio vestibulococlear está inicialmente ligado.
Ilustração mostrando que quase todos os neurônios nos gânglios vestibulares e acústicos são derivados do otocisto como neurônios placodais. Em contraste, todas as células de suporte e Schwann são derivadas das células da crista neural, possivelmente do VII gânglio nervoso ao qual o gânglio vestibulococlear está inicialmente ligado. Vias ascendentes (aferentes ou projetivas) do nervo auditivo
A via ascendente transmite impulsos do órgão espiral (de Corti) para o córtex cerebral (ver imagem a seguir).
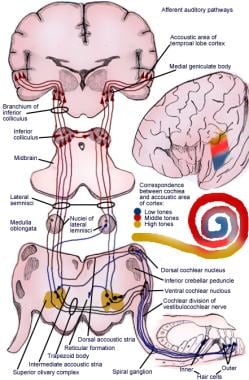 Ilustração da via reflexiva auditiva aferente.
Ilustração da via reflexiva auditiva aferente. Neurônios de primeira ordem do sistema auditivo
As fibras do nervo coclear originam-se de uma agregação de corpos celulares nervosos no gânglio espiral, localizado no modiolo da cóclea. Os neurônios do gânglio espiral são os primeiros de 4 ordens de neurônios entre a cóclea e o cérebro. Eles são células bipolares, porque eles têm 2 conjuntos de processos, ou fibras, que se estendem de extremidades opostas dos corpos celulares. As fibras centrais mais longas, também chamadas de fibras auditivas primárias, formam o nervo coclear, e as fibras periféricas mais curtas estendem-se até as bases das células capilares internas e externas. Elas se estendem radialmente do gânglio espiral até a habenula perforata, uma série de pequenos orifícios sob as células capilares internas. Neste ponto, elas se tornam desmielinizadas para entrar no órgão espiral (de Corti).
Apenas cerca de 30.000 dessas fibras existem, e o maior número delas – cerca de 95% – inervam as células capilares internas. As restantes atravessam o túnel de Corti para interiorizar as células capilares externas. Os processos centrais mais longos dos neurônios cocleares bipolares se unem para formar o tronco do nervo coclear. Estas fibras auditivas primárias saem do modiolo através do meato interno e entram na medula oblonga. A “boca” do canal auditivo interno (CIA) é chamada de porus acusticus. A relação anatômica dos VII e VIII nervos da CIA e a região do ângulo cerebelopontino são áreas anatômicas importantes relacionadas à cirurgia da base do crânio e neuro-otologistas.
Os VII e VIII nervos estão enclausurados em tecido glial ao longo do seu curso intracraniano. Estes nervos estão rodeados por células de Schwann que começam na CIA perto do porus acusticus. A zona de Obersteiner-Redlich é a junção glial-Schwann.
O gânglio Scarpa (vestibular) encontra-se aproximadamente no meio da CIA. A divisão do nervo craniano (CN) VIII em ramos cocleares e vestibulares pode ocorrer no segmento medial da CIA ou no espaço subaracnoideo. A metade posterior da CIA é ocupada pelos ramos superior e inferior do nervo vestibular. O nervo coclear está localizado anteroinferior no canal. O VII nervo está localizado na porção ântero-superior da CIA. Uma crista vertical (barra de Bill) separa os nervos vestibulares facial e superior na parte superior da CIA. A crista transversal separa esta porção superior da CIA da parte inferior contendo as divisões vestibular e coclear inferiores.
O ângulo cerebelopontino é um espaço potencial na fossa craniana posterior. Os seus limites são os seguintes:
-
Anterior: Fossa posterior do osso temporal
-
Posteriormente: Superfície anterior do cerebelo
-
Medialmente: Inferior azeitona
-
Superior: Borda inferior das pons e pedúnculo cerebelar
-
Inferior: A amígdala cerebelar
O nervo trigêmeo é visível superior ao ângulo cerebelopontino, enquanto que o IX, X e XI curso nervos inferior. Outras estruturas importantes dentro do ângulo cerebelopontino incluem a artéria cerebelar inferior anterior (AICA), flóculo e abertura lateral do quarto ventrículo (foramen de Luschka). A artéria labiríntica é geralmente um ramo da AICA e fornece a cóclea e o labirinto.
Após a entrada na medula, as fibras nervosas cocleares seguem para o núcleo coclear. O núcleo coclear consiste de 5 tipos celulares distintos, cada um com características morfológicas e fisiológicas distintas, tais como resposta ao início do estímulo, offset do estímulo e modulação da frequência. O núcleo coclear é dividido nas partes dorsal e ventral. As fibras cocleares dividem-se em 2 feixes principais: Um grupo passa lateral e dorsal para o corpo restiforme; o outro grupo permanece ligeiramente ventral e medial ao corpo restiforme e termina no núcleo coclear ventral. As fibras provenientes das bobinas basais da cóclea terminam na parte dorsal do núcleo coclear dorsal. As fibras provenientes das partes apicais da cóclea terminam na parte ventral do núcleo coclear dorsal e do núcleo ventral. Entretanto, algumas fibras passam para neurônios de ordem mais alta ao longo do caminho antes de sinapse.
Neurônios de segunda, terceira e quarta ordem do sistema auditivo
Os corpos celulares dos neurônios de segunda ordem ficam nos núcleos cocleares dorsal e ventral. Algumas fibras do núcleo coclear ventral passam através da linha média para as células do complexo olivariano superior, enquanto outras fazem conexão com as células olivarianas do mesmo lado. O complexo olivariano superior é considerado o primeiro centro do sistema auditivo ascendente, onde convergem as entradas de ambas as orelhas. Juntas, estas fibras formam o corpo trapezoidal (onde se encontram os neurônios de terceira ordem).
Fibras do núcleo coclear dorsal atravessam a linha média para terminarem nas células dos núcleos dos lemniscus laterais. Lá elas são unidas pelas fibras dos núcleos cocleares ventrais de ambos os lados e do complexo olivariano. Os núcleos auditivos acima do complexo olivariano superior podem ser excitatórios ou inibitórios com inputs de cada orelha. O lemnisco é um trato maior, a maioria das fibras que termina no colículo inferior, o centro auditivo do cérebro médio.
O colículo inferior é um núcleo complexo com pelo menos 18 tipos celulares maiores e pelo menos 5 áreas de especialização. Ele está envolvido em todas as áreas do comportamento auditivo, incluindo sensibilidade diferencial para freqüência e intensidade, sonoridade e audição binaural. Embora algumas fibras possam contornar o colículo e terminar, juntamente com as fibras do colículo, no nível superior seguinte, o corpo geniculado medial do tálamo é onde se encontram os neurônios de quarta ordem. Do corpo geniculado medial, há uma projeção ordenada das fibras para uma porção do córtex do lobo temporal.
Entrada auditiva
O córtex auditivo primário é a primeira região do córtex cerebral a receber entrada auditiva. Em humanos e outros primatas, a área acústica primária no córtex cerebral é o giro temporal transversal superior de Heschl, uma crista no lobo temporal, no lábio inferior da fenda profunda entre os lobos temporal e parietal, conhecida como sulco lateral (fissura Sylvian).
Porque cerca da metade das fibras das vias auditivas atravessam a linha média enquanto outras ascendem do mesmo lado do cérebro, cada orelha é representada tanto no córtex direito como no esquerdo. Por esta razão, mesmo quando a área cortical auditiva de um lado é lesada por trauma ou acidente vascular cerebral, a audição binaural pode ser minimamente afetada.
Percepção do som está associada ao giro temporal superior posterior direito (STG). O giro temporal superior contém várias estruturas importantes do cérebro, incluindo Brodmann 41 e 42, marcando a localização do córtex auditivo primário, a região cortical responsável pela sensação de características básicas do som, como tom e ritmo.
A área de associação auditiva está localizada dentro do lobo temporal do cérebro, em uma área chamada área de Wernicke ou área 22. Esta área, próxima ao sulco cerebral lateral, é uma região importante para o processamento de sinais acústicos para que eles possam ser distinguidos como fala, música ou ruído.
As é comum para conexões talamocorticais, núcleos dentro do corpo geniculado medial que enviam fibras para o córtex auditivo também recebem fibras da mesma área do córtex. Tem sido relatada deficiência auditiva devido a lesão cortical bilateral envolvendo ambas as áreas auditivas, mas é extremamente rara. Entretanto, lesões bilaterais do lubrificante temporal têm demonstrado produzir efeitos abrangentes (surdez cortical, na qual vários comportamentos são afetados, incluindo discriminação de fala, localização do som e detecção de sinais fracos e de curta duração).
A imagem a seguir mostra as vias auditivas consciente e reflexiva aferente.
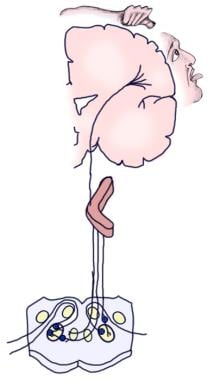 Ilustração da via auditiva consciente.
Ilustração da via auditiva consciente. 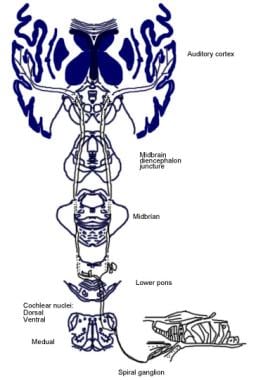 Ilustração da via auditiva reflexiva.
Ilustração da via auditiva reflexiva. Vias descendentes do nervo auditivo
Embora as vias auditivas consciente e reflexiva aferente, também existem vias eferentes descendentes. As projecções eferentes do cérebro para a cóclea também desempenham um papel na percepção do som. Em geral, as vias descendentes têm um efeito inibidor sobre as fibras ascendentes, e tendem a proporcionar auto-regulação ao sistema auditivo. Cada estação auditiva de relé é considerada duplamente valorizada, fornecendo assim um caminho para que os impulsos de entrada sejam influenciados internamente, modificados ou negados.
Paralelo com o caminho ascendente dos núcleos cocleares até o córtex é um caminho descendente do córtex até os núcleos cocleares. Em ambas as vias, algumas das fibras permanecem do mesmo lado, enquanto outras atravessam a linha média até o lado oposto do cérebro. Evidências de uma linha “esporão” ascendendo do núcleo coclear dorsal até o cerebelo e outra descendo do colículo inferior até o cerebelo também existem.
O significado dessas conexões cerebrais não é claro, mas elas podem antever o desenvolvimento evolutivo do córtex cerebral. Do complexo olivariano superior, uma região da medula oblonga, também se origina um trato fibroso chamado feixe olivococlear (veja a imagem abaixo). Ele constitui um sistema eferente, ou loop de retroalimentação, pelo qual os impulsos nervosos, considerados inibidores, atingem as células capilares.
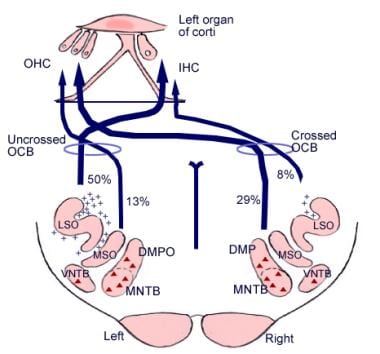 Ilustração das vias descendentes do nervo auditivo. Os cruzamentos representam pequenos neurônios olivococleares (OCB); os triângulos representam grandes neurônios OCB. DMP, DMPO = núcleo pré-olivar medial dorsal; IHC = célula capilar interna; LSO = núcleo olivar superior lateral; MNTB = núcleo olivar medial do corpo trapezoidal; MSO = núcleo olivar superior medial; OHC = célula capilar externa; VNTB = núcleo ventral do corpo trapezoidal.
Ilustração das vias descendentes do nervo auditivo. Os cruzamentos representam pequenos neurônios olivococleares (OCB); os triângulos representam grandes neurônios OCB. DMP, DMPO = núcleo pré-olivar medial dorsal; IHC = célula capilar interna; LSO = núcleo olivar superior lateral; MNTB = núcleo olivar medial do corpo trapezoidal; MSO = núcleo olivar superior medial; OHC = célula capilar externa; VNTB = núcleo ventral do corpo trapezoidal. Anatomia microscópica
Ver o ouvido interno para detalhes sobre a anatomia microscópica e a inervação das células do órgão espiral (de Corti) . As imagens seguintes descrevem esta área.
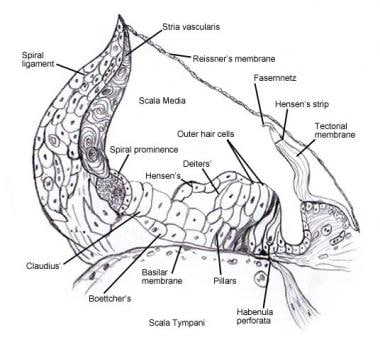 Órgão espiral (de Corti).
Órgão espiral (de Corti).  Micrografia eletrônica de varredura da superfície superior do órgão de Corti após a remoção da membrana tectorial. Existem 3 filas de células capilares externas com a sua característica estereocílios em V. Uma única fila de células capilares internas com uma fila ligeiramente curva de estereocílios também é mostrada.
Micrografia eletrônica de varredura da superfície superior do órgão de Corti após a remoção da membrana tectorial. Existem 3 filas de células capilares externas com a sua característica estereocílios em V. Uma única fila de células capilares internas com uma fila ligeiramente curva de estereocílios também é mostrada. 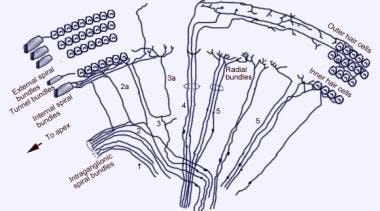 O padrão de inervação aferente da cóclea. A ilustração é mostrada através da membrana vestibular (de Reissner), olhando “para baixo” no órgão espiral (de Corti). Os principais feixes de fibras são 1 e 2; 2a e 3a são fibras espirais internas; 4 são fibras espirais externas; 5 e 6 são fibras radiais que inervam as células capilares internas.
O padrão de inervação aferente da cóclea. A ilustração é mostrada através da membrana vestibular (de Reissner), olhando “para baixo” no órgão espiral (de Corti). Os principais feixes de fibras são 1 e 2; 2a e 3a são fibras espirais internas; 4 são fibras espirais externas; 5 e 6 são fibras radiais que inervam as células capilares internas.