発生学
発生学的には、前庭神経節(音響前庭神経節)は最初第7神経節(顔面神経節)と融合して発生します。 その後、徐々に3つの神経節に分離していきます。 前庭神経節と蝸牛神経節の神経芽細胞はほとんど耳小骨上皮にのみ由来しており、他の頭部感覚神経節では神経節プラコードと神経堤プラコードが神経芽細胞集団に広く寄与しているのと対照的である。 しかし、支持するシュワン細胞とサテライト細胞は、すべての頭蓋神経節と同様に、完全に神経堤由来であり、明らかに顔面神経の神経節から発生している(下の画像参照)。
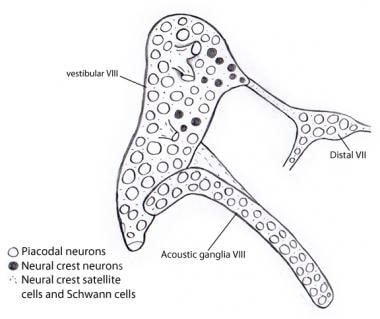 前庭神経節と音響神経節のほとんどすべてのニューロンがプラコダールニューロンとして耳小骨から派生することを示すイラスト。 一方、支持細胞やシュワン細胞はすべて神経堤細胞に由来し、おそらく前庭蝸牛神経節が最初に付着している第VII神経節に由来している。
前庭神経節と音響神経節のほとんどすべてのニューロンがプラコダールニューロンとして耳小骨から派生することを示すイラスト。 一方、支持細胞やシュワン細胞はすべて神経堤細胞に由来し、おそらく前庭蝸牛神経節が最初に付着している第VII神経節に由来している。 聴神経の上行(求心または投射)経路
上行経路は、(コルティ)螺旋器官から大脳皮質にインパルスを伝達する(下図参照)
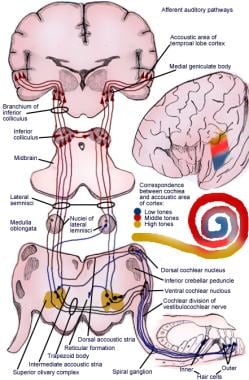 求心性聴覚反射経路の図解。
求心性聴覚反射経路の図解。 聴覚系の一次ニューロン
蝸牛神経の線維は、蝸牛の中隔にある螺旋神経節にある神経細胞体の集合体から発生する。 螺旋神経節は蝸牛と大脳の間にある4次ニューロンのうちの1次ニューロンである。 細胞体の反対側の端から2組の突起(線維)が伸びているため、両極性細胞である。 長い方の中心線維は一次聴覚線維とも呼ばれ、蝸牛神経を形成し、短い方の末梢線維は内毛細胞と外毛細胞の基部まで伸びている。 これらの線維は、螺旋神経節から半径方向に、内毛細胞の下にある一連の小さな穴である穿孔手羽まで伸びている。 この時点で脱髄し、コルチ螺旋器官に入ります。
これらの線維は約3万本しか存在せず、その大部分(約95%)が内毛細胞を神経支配しています。 残りはコルティのトンネルを越えて、外側の有毛細胞を支配している。 両極性蝸牛神経細胞の長い中央突起は、蝸牛神経幹を形成するために一体化されます。 これらの一次聴覚線維は内肉孔を通って中耳を出て、延髄に入る。 内耳道(IAC)の “口 “はporus acusticusと呼ばれます。 第VII、VIII神経は頭蓋内をグリア組織で覆われているため、頭蓋底手術や神経耳鼻科医にとって重要な解剖学的領域である。 これらの神経は、耳介孔に近いIACからシュワン細胞に囲まれている。 Obersteiner-Redlich zoneはグリアとシュワンの接合部です。
スカルパ(前庭)神経節はIACのほぼ中央に位置しています。 頭蓋神経VIIIが蝸牛枝と前庭枝に分岐するのは、IACの内側セグメントかクモ膜下腔である。 IACの後半分は、前庭神経の上枝と下枝が占めています。 蝸牛神経は管内の前内側に位置しています。 第VII神経はIACの前上方に位置しています。 IACの上部で顔面神経と上前壁神経は垂直紋(ビルバー)で隔てられています。 横紋は、IACのこの上部と、下部前庭および蝸牛部門を含む下部とを分離する。
小脳橋角は、後頭蓋窩の潜在的空間である。 その境界は次の通りである:
-
前方。 側頭骨後窩
-
後方:側頭骨後窩。 小脳の前面
-
内側。 下オリーブ
-
上:大脳皮質と小脳小節の下縁
-
下:小脳の下縁
-
下:小脳小節の下縁。 小脳扁桃
小脳橋角の上方に三叉神経が見え、下方に第IX、第X、第XI神経が走行しています。 小脳角内の他の重要な構造としては、前下小脳動脈(AICA)、小丘、第4脳室外側開口部(Luschka孔)などがある。 迷路動脈は通常AICAの枝であり、蝸牛と迷路に供給している。
蝸牛神経線維は髄質に入った後、蝸牛核に進む。 蝸牛核は5種類の細胞からなり、それぞれ刺激オンセット、刺激オフセット、周波数変調に対する反応など、形態的、生理的に異なる特徴を持つ。 蝸牛核は背側と腹側に分かれています。 蝸牛線維は主に2つの束に分かれる。 もう1つの束は、静止体に対してやや腹側で内側を通り、腹側の蝸牛核で終端します。 蝸牛の基底膜から来る線維は、背側蝸牛核の背側で終末を迎えることが分かっている。 蝸牛の先端部からの線維は、背側蝸牛核の腹側部分と腹側核で終末を迎える。 しかし、いくつかの線維は、シナプスする前に経路のさらに上の高次ニューロンに通過する。
聴覚系の2次、3次、4次ニューロン
2次ニューロンの細胞体は、背側および腹側の蝸牛核にある。 腹側蝸牛核からの線維の一部は正中線を越えて上側嗅球複合体の細胞へ、また他の線維は同じ側の嗅球細胞へ接続される。 上唇小体部は、両耳からの入力が収束する上行性聴覚系の最初の中枢と考えられている。 8363>
背側蝸牛核からの線維は、正中線を横切って外側レムニスカスの核の細胞上で終止する。 そこで両側の腹側蝸牛核からの線維とオリーブ複合体からの線維とが合流する。 上オリーブ複合体より上の聴覚核は、それぞれの耳からの入力で興奮性にも抑制性にもなりうる。 下小丘は、少なくとも18の主要な細胞型と少なくとも5つの特殊化領域を持つ複雑な核であり、その繊維のほとんどは、中脳の聴覚中枢である下小丘で終わる。 下小丘は、周波数と強度の差、ラウドネス、両耳聴など、聴覚行動のあらゆる領域に関与している。 一部の線維は小丘を迂回して、小丘からの線維とともに次の高次のレベルで終わることもあるが、視床の内側被蓋体は第4次ニューロンが存在する場所である。 内側性器体からは、側頭葉の皮質の一部に繊維が整然と投射されている。
聴覚入力
一次聴覚皮質は、大脳皮質の中で最初に聴覚入力を受ける領域である。 ヒトや他の霊長類では、大脳皮質の一次音響領域は、側溝(シルビウス裂)として知られる側頭葉と頭頂葉の間の深い裂け目の下唇にある側頭葉の隆起であるヘシュルの上横側頭回である。
聴覚経路の繊維の約半分は正中線を横切り、他の繊維は脳の同じ側を上昇するため、それぞれの耳は右と左の両方の皮質に表現されます。 このため、外傷や脳卒中などで片側の聴覚皮質が損傷しても、両耳の聴覚にはほとんど影響がない。
音の知覚は右後方の上側頭回(STG)と関連している。 上側頭回にはBrodmann 41と42を含む脳のいくつかの重要な構造があり、ピッチやリズムといった音の基本的な特性の感覚を担う皮質領域である一次聴覚皮質の位置を示す。
聴覚連合野は脳の側頭葉内のWernicke領域または22領域と呼ばれる領域にある。 この領域は外側脳溝の近くにあり、音響信号を処理して音声、音楽、雑音として区別するのに重要な領域である。
視床皮質接続によく見られるように、聴覚皮質に線維を送る内側被蓋体内の核は、皮質の同じ領域から線維を受け取ることもある。 両耳の皮質損傷による聴覚障害は報告されているが、極めて稀である。 しかし、側頭葉の両側性病変は広範囲の影響(音声弁別、音の定位、微弱で持続時間の短い信号の検出など、いくつかの行動が影響を受ける皮質性難聴)をもたらすことが示されている
意識的聴覚経路と反射的求心性聴覚経路を以下の画像で説明する。
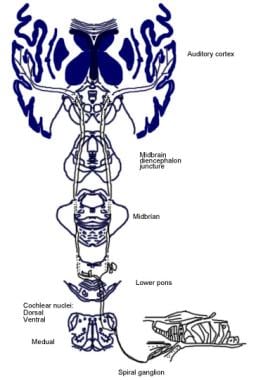 反射性聴覚路の図解。
反射性聴覚路の図解。 聴神経の下行性経路
意識的・反射的な求心性の聴覚経路のほかに、下行性の求心性経路も存在する。 脳から蝸牛への遠心性投射もまた、音の知覚に関与している。 一般に、下行性経路は上行性線維を抑制する効果があり、聴覚系に自己調節を与える傾向がある。 蝸牛核から大脳皮質に上る経路と平行して、大脳皮質から蝸牛核に下る経路がある。 どちらの経路も、同じ側に残る繊維もあれば、正中線を横切って反対側に出る繊維もある。 また、背側蝸牛核から小脳に向かう “spur “線と下小丘から小脳に向かう “spur “線も存在する。
これらの大脳結合の意味は明らかではないが、大脳皮質の進化的発達に先行している可能性がある。 また、延髄の上オリーブ複合体という部位から、オリーブ束という繊維路が発生している(下図参照)。 8363>
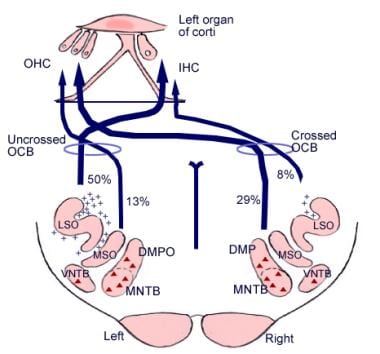 聴神経の下行性経路の図解。 十字は小脳蝸牛束(OCB)ニューロン、三角は大脳蝸牛束(OCB)ニューロンを表す。 DMP、DMPO=背内側前葉核、IHC=内毛細胞、LSO=外側上葉核、MNTB=台形体内側核、MSO=内側上葉核、OHC=外毛細胞、VNTB=台形体腹側核。
聴神経の下行性経路の図解。 十字は小脳蝸牛束(OCB)ニューロン、三角は大脳蝸牛束(OCB)ニューロンを表す。 DMP、DMPO=背内側前葉核、IHC=内毛細胞、LSO=外側上葉核、MNTB=台形体内側核、MSO=内側上葉核、OHC=外毛細胞、VNTB=台形体腹側核。 ミクロの解剖学
螺旋器官(コルティ)の細胞のミクロの解剖学と神経支配に関する詳細は、内耳を参照してください。 8363>
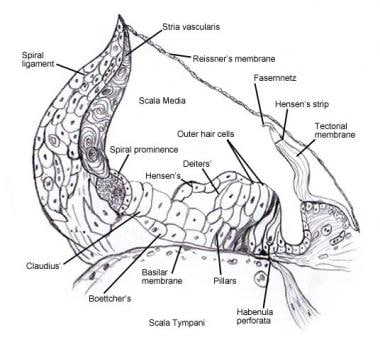 スパイラル器官(Corti)。
スパイラル器官(Corti)。  胸膜を除去したコルチ器官の上面の走査型電子顕微鏡写真。 3列の外有毛細胞があり、特徴的なV字型の立体繊毛が並んでいる。 また、わずかに湾曲した定位繊毛列を持つ1列の内毛細胞も示されている。
胸膜を除去したコルチ器官の上面の走査型電子顕微鏡写真。 3列の外有毛細胞があり、特徴的なV字型の立体繊毛が並んでいる。 また、わずかに湾曲した定位繊毛列を持つ1列の内毛細胞も示されている。 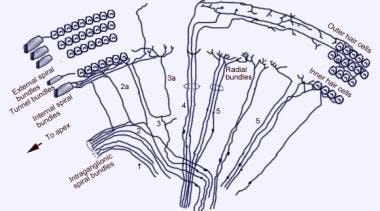 蝸牛の求心性神経支配パターン。 前庭(ライスナー)膜を通して、(コルティ)螺旋器官を「見下ろす」形で図示している。 主な繊維束は1と2、2aと3aは内螺旋繊維、4は外螺旋繊維、5と6は内有毛細胞を支配する放射状繊維である。
蝸牛の求心性神経支配パターン。 前庭(ライスナー)膜を通して、(コルティ)螺旋器官を「見下ろす」形で図示している。 主な繊維束は1と2、2aと3aは内螺旋繊維、4は外螺旋繊維、5と6は内有毛細胞を支配する放射状繊維である。