Embryologi
Embryologisk set udvikles det vestibulocochleære (akustisk-vestibulære) ganglion oprindeligt sammenvokset med den VII. Adskillelsen af de 3 komponenter sker gradvist. Neuroblasterne i de vestibulære og cochleære (akustiske) ganglier stammer næsten udelukkende fra otocystepithelet, i modsætning til andre kraniale sensoriske ganglier, hvor både ganglionære og neuralkystplacoder yder et omfattende bidrag til neuroblastepopulationerne. De understøttende Schwann- og satellitceller er imidlertid, som i alle kraniale ganglier, udelukkende af neuralkammen og stammer tilsyneladende fra ganglien af ansigtsnerven (se billedet nedenfor).
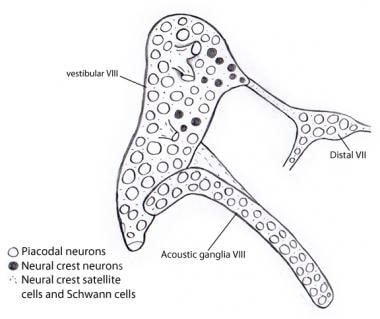 Illustration, der viser, at næsten alle neuroner i de vestibulære og akustiske ganglier er afledt fra otocysten som placodale neuroner. Derimod stammer alle støtte- og Schwann-cellerne fra neurale kamceller, muligvis fra det VII. nerveganglion, som det vestibulocochleære ganglion oprindeligt er knyttet til.
Illustration, der viser, at næsten alle neuroner i de vestibulære og akustiske ganglier er afledt fra otocysten som placodale neuroner. Derimod stammer alle støtte- og Schwann-cellerne fra neurale kamceller, muligvis fra det VII. nerveganglion, som det vestibulocochleære ganglion oprindeligt er knyttet til. Afferente (afferente eller projektive) baner i hørenerven
Den opstigende bane overfører impulser fra spiralorganet (af Corti) til hjernebarken (se følgende billede).
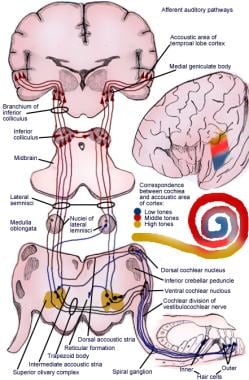 Illustration af den afferente hørerefleksbane.
Illustration af den afferente hørerefleksbane. Neuroner af første orden i det auditive system
Fibrene i den cochleare nerve stammer fra en samling af nervecellelegemer i det spiralformede ganglion, der er placeret i modiolus i cochlea. Neuronerne i det spiralformede ganglion er de første af 4 ordensneuroner mellem cochlea og cerebrum. De er bipolære celler, fordi de har 2 sæt processer, eller fibre, der udgår fra modsatte ender af cellelegemerne. De længere centrale fibre, også kaldet de primære hørefibre, danner den cochleare nerve, og de kortere, perifere fibre strækker sig til baserne af de indre og ydre hårceller. De strækker sig radialt fra det spiralformede ganglion til habenula perforata, en række små huller under de indre hårceller. Her bliver de demyeliniserede for at gå ind i spiralorganet (af Corti).
Der findes kun ca. 30.000 af disse fibre, og størstedelen af dem – ca. 95 % – innerverer de indre hårceller. De resterende krydser Cortis tunnel for at innervere de ydre hårceller. De længere centrale processer af de bipolære cochleare neuroner forenes til den cochleare nervestamme. Disse primære hørefibre forlader modiolus gennem den indre meatus og går ind i medulla oblongata. “Munden” i den indre øregang (IAC) kaldes porus acusticus. Det anatomiske forhold mellem den VII. og VIII. nerve i IAC og cerebellopontine vinkelregionen er vigtige anatomiske områder i forbindelse med kraniebasiskirurgi og neuro-otologer.
Den VII. og VIII. nerve er omsluttet af gliavæv i hele deres intrakranielle forløb. Disse nerver er omgivet af Schwann-celler, der begynder i IAC tæt ved porus acusticus. Obersteiner-Redlich-zonen er glial-Schwann-knudepunktet.
Den Scarpa (vestibulære) ganglion ligger omtrent midt i IAC. Delingen af kranienerve (CN) VIII i de cochleære og vestibulære grene kan ske i det mediale segment af IAC eller i subaraknoidalrummet. Den bageste halvdel af IAC er optaget af den øverste og nederste gren af nervus vestibularis. Den cochleære nerve er placeret anteroinferior i kanalen. Den VII. nerve er placeret i den anterosuperior del af IAC. En lodret kam (Bill bar) adskiller facialisnerven og den overlegne vestibulærnerve i den øverste del af IAC. Den tværgående kam adskiller denne øvre del af IAC fra den nedre del, der indeholder de nedre vestibulære og cochleære afdelinger.
Den cerebellopontine vinkel er et potentielt rum i den bageste kraniefossa. Dens grænser er som følger:
-
Anteriort: Bagtil: Fossa temporalis posteriore
-
Posterior: Bagtil: Den bagvedliggende fossa i temporalknoglen
-
Forreste overflade af lillehjernen
-
Medialt: Inferior olive
-
Overst: Inferior grænse af pons og cerebellar peduncle
-
Inferior: Cerebellar tonsil
Nervus trigeminus er synlig over for den cerebellopontine vinkel, mens den IX., X. og XI. nerve løber inferiort. Andre vigtige strukturer i den cerebellopontine vinkel omfatter den forreste inferior cerebellare arterie (AICA), flocculus og den laterale åbning af den fjerde ventrikel (Luschka-formamen). Den labyrintiske arterie er normalt en gren af AICA og forsyner cochlea og labyrinten.
Når de cochleale nervefibre kommer ind i medulla, fortsætter de cochleale nervefibre til den cochleale nucleus. Den cochleære kerne består af 5 forskellige celletyper, som hver især har forskellige morfologiske og fysiologiske egenskaber, såsom respons på stimulusstart, stimulusoffset og frekvensmodulation. Den cochleare kerne er opdelt i en dorsal og en ventral del. De cochleære fibre deler sig i 2 hovedbundter: Den ene gruppe passerer lateralt og dorsalt til det restiformiske legeme; den anden gruppe forbliver lidt ventral og medial til det restiformiske legeme og ender i den ventrale cochleare kerne. Fibre, der kommer fra de basale spoler i cochlea, er blevet fundet at ende i den dorsale del af den dorsale cochlear nucleus. Fibrene fra de apikale dele af cochlea ender i den ventrale del af den dorsale cochlear nucleus og i den ventrale nucleus. Nogle fibre passerer dog til neuroner af højere orden længere fremme i banen, før de synapserer.
Neuroner af anden, tredje og fjerde orden i det auditive system
Cellekroppene af neuronerne af anden orden ligger i den dorsale og ventrale cochleare kerne. Nogle fibre fra den ventrale cochleære kerne passerer over midterlinjen til cellerne i det overlegne olivariekompleks, mens andre laver forbindelse med olivariecellerne i samme side. Det overlegne olivariekompleks anses for at være det første center i det opstigende auditive system, hvor input fra begge ører konvergerer. Tilsammen danner disse fibre trapezlegemet (hvor tredjeordensneuronerne er placeret).
Fibre fra den dorsale cochlear nucleus krydser midterlinjen for at ende på cellerne i kernerne i den laterale lemniscus. Der forenes de med fibrene fra de ventrale cochleære kerner i begge sider og fra olivariekomplekset. Auditive kerner over det overlegne olivariekompleks kan være excitatoriske eller inhibitoriske med input fra hvert øre. Lemniscus er et vigtigt tractus, hvoraf de fleste af fibrene ender i inferior colliculus, det auditive center i mellemhjernen.
Inferior colliculus er en kompleks kerne med mindst 18 hovedcelletyper og mindst 5 specialiseringsområder. Den er involveret i alle områder af auditiv adfærd, herunder differentiel følsomhed for frekvens og intensitet, lydstyrke og binaural hørelse. Selv om nogle fibre kan gå uden om colliculus og sammen med fibrene fra colliculus ende på det næste højere niveau, er det thalamus’ mediale genikulære legeme, hvor neuronerne af fjerde orden er placeret. Fra det mediale geniculaturlegeme er der en ordnet projektion af fibre til en del af cortexen i tindingelappen.
Auditivt input
Den primære auditive cortex er den første region i hjernebarken, der modtager auditivt input. Hos mennesker og andre primater er det primære akustiske område i hjernebarken Heschls overlegne transversale temporale gyri, en højderyg i tindingelappen, på underlæben af den dybe kløft mellem tindingelappen og parietallappen, kendt som den laterale sulcus (Sylvian fissur).
Da omkring halvdelen af fibrene i hørebanerne krydser midterlinjen, mens andre stiger op på samme side af hjernen, er hvert øre repræsenteret i både højre og venstre cortex. Af denne grund kan binaural hørelse være minimalt påvirket, selv når det auditive kortikale område i den ene side er skadet ved traume eller slagtilfælde.
Lydopfattelse er forbundet med den højre posterior superior temporal gyrus (STG). Den overlegne tidsmæssige gyrus indeholder flere vigtige strukturer i hjernen, herunder Brodmann 41 og 42, der markerer placeringen af den primære auditive cortex, den kortikale region, der er ansvarlig for fornemmelsen af grundlæggende lydkarakteristika såsom tonehøjde og rytme.
Det auditive associationsområde er placeret i hjernens tindingelap, i et område kaldet Wernicke-området eller område 22. Dette område, nær den laterale cerebrale sulcus, er et vigtigt område til behandling af akustiske signaler, så de kan skelnes som tale, musik eller støj.
Som det er almindeligt for thalamokortikale forbindelser, modtager kerner i det mediale geniculære legeme, der sender fibre til den auditive cortex, også fibre fra det samme område af cortex. Der er rapporteret om nedsat hørelse som følge af bilateral kortikal skade, der involverer begge auditive områder, men det er yderst sjældent. Det er imidlertid vist, at bilaterale læsioner af den temporale lube har vist sig at give vidtrækkende virkninger (kortikal døvhed, hvor flere adfærdsmønstre påvirkes, herunder talediskrimination, lokalisering af lyd og detektion af svage, kortvarige signaler).
Det følgende billede skildrer de bevidste og refleksafferente auditive baner.
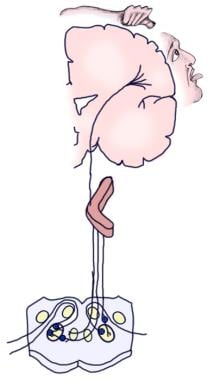 Illustration af den bevidste auditive bane.
Illustration af den bevidste auditive bane. 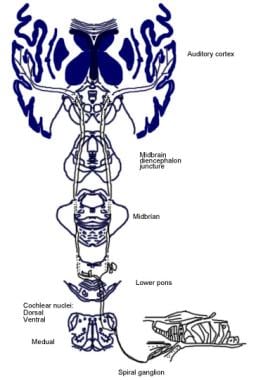 Illustration af den auditive refleksbane.
Illustration af den auditive refleksbane. Audioriskenes nedadgående baner
Udover de bevidste og refleksafferente auditive baner findes der også nedadgående efferente baner. Efferente projektioner fra hjernen til cochlea spiller også en rolle i opfattelsen af lyd. Generelt har de nedadgående veje en hæmmende virkning på de opadgående fibre, og de har en tendens til at give selvregulering til det auditive system. Hver relæauditiv station anses for at være dobbelt innerveret, hvilket giver mulighed for, at indkommende impulser kan påvirkes, modificeres eller negeres internt.
Parallelt med den opstigende vej fra de cochleære kerner til cortexen er der en nedstigende vej fra cortexen til de cochleære kerner. I begge baner forbliver nogle af fibrene på den samme side, mens andre krydser midterlinjen til den modsatte side af hjernen. Der findes også beviser for en “spore”-linje, der går opad fra den dorsale cochlear kerne til lillehjernen, og en anden, der går nedad fra inferior colliculus til lillehjernen.
Den betydning, som disse cerebrale forbindelser har, er ikke klar, men de kan gå forud for den evolutionære udvikling af hjernebarken. Fra det overlegne olivariekompleks, et område i medulla oblongata, udgår også en fiberbane kaldet olivocochlear bundle (se billedet nedenfor). Den udgør et efferent system, eller feedback loop, hvorigennem nerveimpulser, der menes at være hæmmende, når hårcellerne.
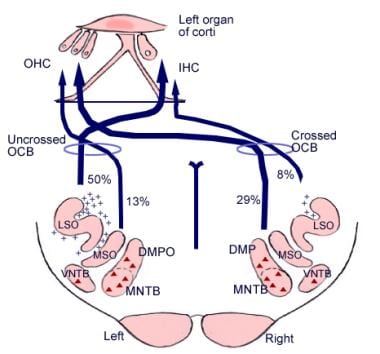 Illustration af de nedadgående baner i hørenerven. Krydsene repræsenterer små olivocochlear bundle (OCB)-neuroner; trekanterne repræsenterer store OCB-neuroner. DMP, DMPO = dorsal medial preolivær kerne; IHC = indre hårcelle; LSO = lateral superior olivary nucleus; MNTB = medial nucleus of trapezoid body; MSO = medial superior olivary nucleus; OHC = ydre hårcelle; VNTB = ventral nucleus of trapezoid body.
Illustration af de nedadgående baner i hørenerven. Krydsene repræsenterer små olivocochlear bundle (OCB)-neuroner; trekanterne repræsenterer store OCB-neuroner. DMP, DMPO = dorsal medial preolivær kerne; IHC = indre hårcelle; LSO = lateral superior olivary nucleus; MNTB = medial nucleus of trapezoid body; MSO = medial superior olivary nucleus; OHC = ydre hårcelle; VNTB = ventral nucleus of trapezoid body. Mikroskopisk anatomi
Se Indre øre for nærmere oplysninger om den mikroskopiske anatomi og innervation af cellerne i spiralorganet (Corti) . De følgende billeder viser dette område.
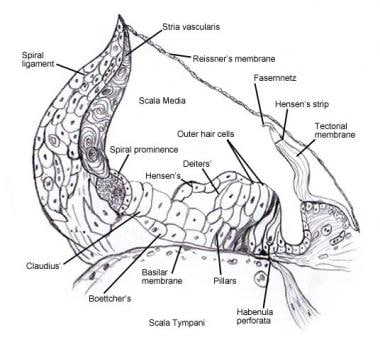 Spiralorgan (of Corti).
Spiralorgan (of Corti).  Scanningelektronmikroskopisk billede af den øvre overflade af Corti-organet efter fjernelse af tectorialmembranen. Der er 3 rækker af ydre hårceller med deres karakteristiske V-arrangement af stereocilia. En enkelt række indre hårceller med en let buet række af stereocilia er også vist.
Scanningelektronmikroskopisk billede af den øvre overflade af Corti-organet efter fjernelse af tectorialmembranen. Der er 3 rækker af ydre hårceller med deres karakteristiske V-arrangement af stereocilia. En enkelt række indre hårceller med en let buet række af stereocilia er også vist. 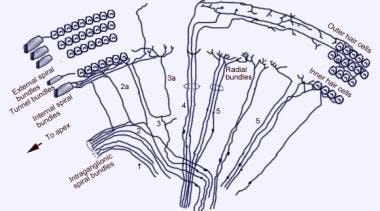 Cochleaens afferente innervationsmønster. Illustrationen er vist gennem den vestibulære (Reissners) membran, idet man ser “ned” på spiralorganet (Corti-organet). De vigtigste fiberbundter er 1 og 2; 2a og 3a er indre spiralfibre; 4 er ydre spiralfibre; 5 og 6 er radiale fibre, der innerverer de indre hårceller.
Cochleaens afferente innervationsmønster. Illustrationen er vist gennem den vestibulære (Reissners) membran, idet man ser “ned” på spiralorganet (Corti-organet). De vigtigste fiberbundter er 1 og 2; 2a og 3a er indre spiralfibre; 4 er ydre spiralfibre; 5 og 6 er radiale fibre, der innerverer de indre hårceller.