Embryologie
Embryologisch ontwikkelt het vestibulocochleair (acousticovestibulair) ganglion zich aanvankelijk versmolten met het VIIe zenuw (gezichts) ganglion. De scheiding van de 3 componenten gebeurt geleidelijk. De vestibulaire en cochleaire (akoestische) ganglia neuroblasten zijn bijna uitsluitend afkomstig van het otocyste epitheel, in tegenstelling tot andere craniale sensorische ganglia waar zowel ganglionische als neurale crest placodes uitgebreide bijdragen leveren aan de neuroblastpopulaties. De ondersteunende Schwann- en satellietcellen zijn echter, net als in alle craniale ganglia, geheel van neurale- crest oorsprong, kennelijk voortkomend uit het ganglion van de nervus facialis (zie de afbeelding hieronder).
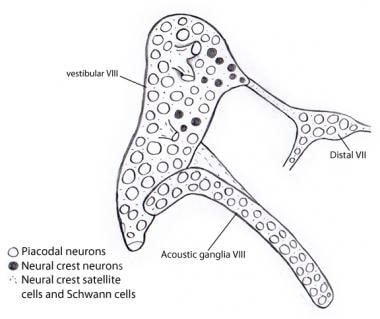 Illustratie die laat zien dat bijna alle neuronen in de vestibulaire en akoestische ganglia als placodale neuronen afkomstig zijn uit de otocyst. Daarentegen zijn alle steun- en Schwanncellen afgeleid van neurale crestcellen, mogelijk van het VIIe zenuwganglion waaraan het vestibulocochleair ganglion aanvankelijk is gehecht.
Illustratie die laat zien dat bijna alle neuronen in de vestibulaire en akoestische ganglia als placodale neuronen afkomstig zijn uit de otocyst. Daarentegen zijn alle steun- en Schwanncellen afgeleid van neurale crestcellen, mogelijk van het VIIe zenuwganglion waaraan het vestibulocochleair ganglion aanvankelijk is gehecht. Ascenderende (afferente of projectieve) banen van de gehoorzenuw
De ascenderende baan zendt impulsen van het spiraalvormige orgaan (van Corti) naar de cerebrale cortex (zie de volgende afbeelding).
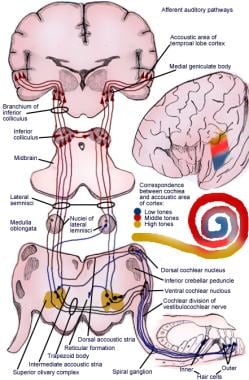 Illustratie van de afferente auditieve reflexbaan.
Illustratie van de afferente auditieve reflexbaan. Neuronen van de eerste orde van het auditieve systeem
De vezels van de cochleaire zenuw zijn afkomstig van een aggregatie van zenuwcellichamen in het spiraalvormige ganglion, gelegen in de modiolus van het slakkenhuis. De neuronen van het spiraal ganglion zijn de eerste van de 4 orde neuronen tussen het slakkenhuis en de kleine hersenen. Het zijn bipolaire cellen, omdat zij 2 sets uitsteeksels, of vezels, hebben die uit tegenovergestelde uiteinden van de cellichamen komen. De langere centrale vezels, ook wel de primaire gehoorvezels genoemd, vormen de cochleaire zenuw, en de kortere, perifere vezels strekken zich uit tot de bases van de binnenste en buitenste haarcellen. Zij strekken zich radiaal uit van het spiraal ganglion tot aan de habenula perforata, een reeks kleine gaatjes onder de binnenste haarcellen. Op dit punt worden zij gedemyeliniseerd om het spiraalvormige orgaan (van Corti) binnen te gaan.
Er bestaan slechts ongeveer 30.000 van deze vezels, en het grootste aantal daarvan – ongeveer 95% – bedienen de binnenste haarcellen. De rest kruist de tunnel van Corti om de buitenste haarcellen te innerveren. De langere centrale processen van de bipolaire cochleaire neuronen verenigen zich om de cochleaire zenuwstam te vormen. Deze primaire auditieve vezels verlaten de modiolus via de interne meatus en gaan de medulla oblongata binnen. De “mond” van de interne gehoorgang (IAC) wordt de porus acusticus genoemd. De anatomische relatie van de VIIe en VIIIe zenuwen in de IAC en de cerebellopontine hoek regio zijn belangrijke anatomische gebieden met betrekking tot schedelbasis chirurgie en neuro-otologen.
De VIIe en VIIIe zenuwen zijn omhuld door gliaal weefsel gedurende hun gehele intracraniële traject. Deze zenuwen zijn omgeven door Schwann cellen beginnend in het IAC dicht bij de porus acusticus. De Obersteiner-Redlich-zone is de gliaal-Schwann-verbinding.
Het Scarpa (vestibulair) ganglion ligt ongeveer in het midden van het IAC. De splitsing van de hersenzenuw (CN) VIII in de cochleaire en vestibulaire takken kan plaatsvinden in het mediale segment van het IAC of in de subarachnoïdale ruimte. De achterste helft van de IAC wordt ingenomen door de superieure en inferieure takken van de nervus vestibularis. De n. cochlea ligt antero-inferior in het kanaal. De n. VIIe bevindt zich in het anterosuperiore gedeelte van de IAC. Een verticale kam (Bill bar) scheidt de nervus facialis en de nervus vestibularis superior in het bovenste deel van het IAC. De transversale kam scheidt dit bovenste deel van het IAC van het onderste deel dat de lagere vestibulaire en cochleaire divisies bevat.
De cerebellopontine hoek is een potentiële ruimte in de achterste schedelgroeve. De grenzen ervan zijn als volgt:
-
Anterior: Achterste fossa van het slaapbeen
-
Posteriorly: Anterior surface of the cerebellum
-
Mediaal: Inferior olive
-
Superiorly: Inferior border of the pons and cerebellar peduncle
-
Inferiorly: De cerebellaire tonsil
De nervus trigeminus is zichtbaar superieur aan de cerebellopontine hoek, terwijl de IXe, Xe, en XIe zenuwen inferieur lopen. Andere belangrijke structuren binnen de cerebellopontine hoek zijn de anterieure inferieure cerebellaire slagader (AICA), de flocculus, en de laterale opening van de vierde ventrikel (foramen van Luschka). De labyrintische slagader is gewoonlijk een tak van de AICA en voorziet het slakkenhuis en het labyrint.
Na het medulla te zijn binnengegaan, gaan de cochleaire zenuwvezels naar de cochleaire kern. De cochleaire kern bestaat uit 5 verschillende celtypes, elk met verschillende morfologische en fysiologische kenmerken, zoals respons op stimulus begin, stimulus offset, en frequentie modulatie. De cochleaire kern is verdeeld in een dorsaal en ventraal deel. De cochleaire vezels verdelen zich in 2 hoofdbundels: Eén groep passeert lateraal en dorsaal naar het restiforme lichaam; de andere groep blijft iets ventraal en mediaal van het restiforme lichaam en eindigt in de ventrale cochleaire nucleus. Vezels afkomstig van de basale spoelen van het slakkenhuis bleken te eindigen in het dorsale deel van de dorsale cochleaire kern. De vezels uit de apicale delen van het slakkenhuis eindigen in het ventrale deel van de dorsale cochleaire kern en de ventrale nucleus. Sommige vezels gaan echter naar neuronen van een hogere orde verderop in het pad voordat ze synapsen.
Neuronen van de tweede, derde en vierde orde van het auditieve systeem
De cellichamen van de neuronen van de tweede orde liggen in de dorsale en ventrale cochleaire kernen. Sommige vezels van de ventrale cochleaire kern gaan over de middellijn naar de cellen van het superieure olivariumcomplex, terwijl andere verbinding maken met de olivariumcellen van dezelfde zijde. Het superieure olivariumcomplex wordt beschouwd als het eerste centrum in het opgaande auditieve systeem, waar de inputs van beide oren samenkomen. Samen vormen deze vezels het trapeziumlichaam (waar de neuronen van de derde orde zich bevinden).
Zwembers uit de dorsale cochleaire kern kruisen de middellijn om te eindigen op de cellen van de kernen van de laterale lemniscus. Daar worden zij verbonden door de vezels van de ventrale cochleaire kernen van beide zijden en van het olivariumcomplex. De auditieve kernen boven het superieure olivariumcomplex kunnen excitatoir of inhibitoir zijn met inputs vanuit elk oor. De lemniscus is een belangrijk kanaal, waarvan de meeste vezels eindigen in de inferieure colliculus, het auditieve centrum van de middenhersenen.
De inferieure colliculus is een complexe kern met ten minste 18 belangrijke celtypen en ten minste 5 gebieden van specialisatie. Hij is betrokken bij alle gebieden van het auditieve gedrag, met inbegrip van differentiële gevoeligheid voor frequentie en intensiteit, luidheid, en binauraal horen. Hoewel sommige vezels de colliculus omzeilen en samen met de vezels van de colliculus op een hoger niveau eindigen, is het mediale geniculaatlichaam van de thalamus de plaats waar de neuronen van de vierde orde zich bevinden. Vanuit het mediale geniculate lichaam is er een ordelijke projectie van vezels naar een deel van de cortex van de temporale kwab.
Auditieve input
De primaire auditieve cortex is het eerste gebied van de cerebrale cortex dat auditieve input ontvangt. Bij de mens en andere primaten is het primaire akoestische gebied in de hersenschors de superieure transversale temporale gyri van Heschl, een richel in de temporale kwab, op de onderste lip van de diepe kloof tussen de temporale en pariëtale kwabben, bekend als de laterale sulcus (Sylviaanse spleet).
Omdat ongeveer de helft van de vezels van de auditieve paden de middellijn kruisen, terwijl andere aan dezelfde kant van de hersenen omhoog gaan, is elk oor vertegenwoordigd in zowel de rechter als de linker cortex. Om deze reden, zelfs wanneer het auditieve corticale gebied van één kant door trauma of beroerte wordt verwond, kan het binaurale horen minimaal worden beïnvloed.
De perceptie van geluid wordt geassocieerd met de rechter posterieure superieure temporale gyrus (STG). De superieure temporale gyrus bevat verschillende belangrijke structuren van de hersenen, waaronder Brodmann 41 en 42, die de locatie van de primaire auditieve cortex markeren, het corticale gebied dat verantwoordelijk is voor de sensatie van basiskenmerken van geluid, zoals toonhoogte en ritme.
Het auditieve associatiegebied bevindt zich in de temporale kwab van de hersenen, in een gebied dat het Wernicke-gebied of gebied 22 wordt genoemd. Dit gebied, in de buurt van de laterale sulcus cerebralis, is een belangrijk gebied voor de verwerking van akoestische signalen, zodat ze kunnen worden onderscheiden als spraak, muziek of lawaai.
Zoals gebruikelijk is voor thalamocorticale verbindingen, ontvangen kernen binnen het mediale geniculate lichaam die vezels naar de auditieve cortex sturen, ook vezels uit hetzelfde gebied van de cortex. Slechthorendheid als gevolg van bilateraal corticaal letsel waarbij beide auditieve gebieden betrokken zijn, is gerapporteerd, maar het is uiterst zeldzaam. Er is echter aangetoond dat bilaterale laesies van de temporale lube verstrekkende gevolgen hebben (corticale doofheid, waarbij verschillende gedragingen worden beïnvloed, met inbegrip van spraakdiscriminatie, lokalisatie van geluid, en de detectie van zwakke, kortdurende signalen).
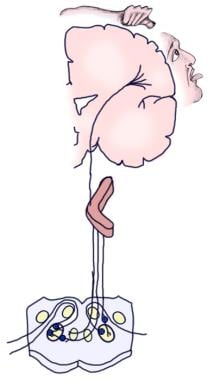
De volgende afbeelding toont de bewuste en reflex afferente auditieve pathways.
 Illustratie van de bewuste auditieve pathway.
Illustratie van de bewuste auditieve pathway. 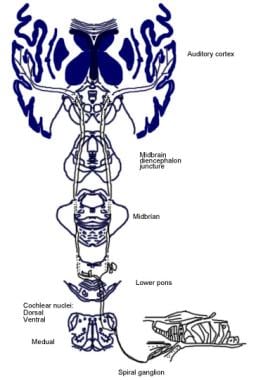 Illustratie van de auditieve reflex pathway.
Illustratie van de auditieve reflex pathway. Dalende banen van de gehoorzenuw
Naast de bewuste en reflexmatige afferente auditieve banen, bestaan er ook dalende efferente banen. Efferente projecties van de hersenen naar het slakkenhuis spelen ook een rol bij de perceptie van geluid. In het algemeen hebben de afgaande paden een remmend effect op de opgaande vezels en zorgen zij voor zelfregulering van het auditieve systeem. Elk auditief relaisstation wordt geacht dubbel geïnnerveerd te zijn, waardoor inkomende impulsen intern kunnen worden beïnvloed, gewijzigd of genegeerd.
Parallel aan de route die van de cochleaire kernen naar de cortex loopt, loopt een route die van de cortex naar de cochleaire kernen loopt. In beide banen blijven sommige vezels aan dezelfde kant, terwijl andere de middellijn oversteken naar de andere kant van de hersenen. Er zijn ook aanwijzingen van een “uitloper” lijn die opklimt van de dorsale cochleaire kern naar het cerebellum en een andere die afdaalt van de inferieure colliculus naar het cerebellum.
De betekenis van deze cerebrale verbindingen is niet duidelijk, maar zij kunnen voorafgaan aan de evolutionaire ontwikkeling van de cerebrale cortex. Uit het superieure olivariumcomplex, een gebied in de medulla oblongata, ontspringt ook een vezelkanaal dat de olivocochleaire bundel wordt genoemd (zie de afbeelding hieronder). Deze vormt een efferent systeem, of terugkoppellus, waardoor zenuwimpulsen, die geacht worden remmend te werken, de haarcellen bereiken.
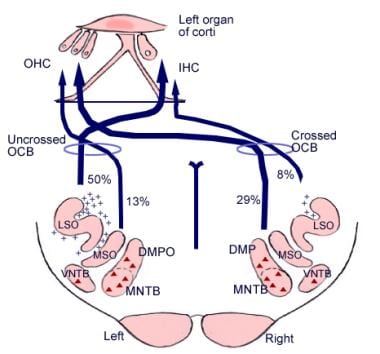 Illustratie van de afgaande paden van de gehoorzenuw. De kruisjes stellen kleine olivocochleaire bundel (OCB) neuronen voor; de driehoekjes stellen grote OCB neuronen voor. DMP, DMPO = dorsale mediale preolivaire kern; IHC = binnenste haarcel; LSO = laterale superieure olivary nucleus; MNTB = mediale kern van trapezium lichaam; MSO = mediale superieure olivary kern; OHC = buitenste haarcel; VNTB = ventrale kern van trapezium lichaam.
Illustratie van de afgaande paden van de gehoorzenuw. De kruisjes stellen kleine olivocochleaire bundel (OCB) neuronen voor; de driehoekjes stellen grote OCB neuronen voor. DMP, DMPO = dorsale mediale preolivaire kern; IHC = binnenste haarcel; LSO = laterale superieure olivary nucleus; MNTB = mediale kern van trapezium lichaam; MSO = mediale superieure olivary kern; OHC = buitenste haarcel; VNTB = ventrale kern van trapezium lichaam. Microscopische anatomie
Zie Binnenoor voor bijzonderheden betreffende de microscopische anatomie en de innervatie van de cellen in het spiraalvormige orgaan (van Corti) . De volgende afbeeldingen tonen dit gebied.
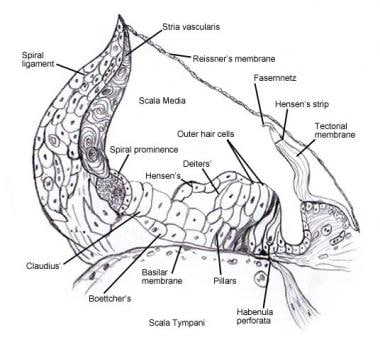 Spiraalvormig orgaan (van Corti).
Spiraalvormig orgaan (van Corti).  Scanning electronen microfoto van het bovenoppervlak van het orgaan van Corti na verwijdering van het tectoriaal membraan. Er zijn 3 rijen buitenste haarcellen met hun karakteristieke V-geordende stereocilia. Een enkele rij van binnenste haarcellen met een licht gebogen rij stereocilia is ook getoond.
Scanning electronen microfoto van het bovenoppervlak van het orgaan van Corti na verwijdering van het tectoriaal membraan. Er zijn 3 rijen buitenste haarcellen met hun karakteristieke V-geordende stereocilia. Een enkele rij van binnenste haarcellen met een licht gebogen rij stereocilia is ook getoond. 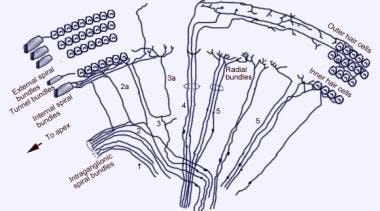 Het afferente innervatiepatroon van het slakkenhuis. De illustratie wordt getoond door het vestibulaire (Reissner’s) membraan, naar “beneden” kijkend op het spiraalvormige orgaan (van Corti). De voornaamste vezelbundels zijn 1 en 2; 2a en 3a zijn interne spiraalvormige vezels; 4 zijn externe spiraalvormige vezels; 5 en 6 zijn radiale vezels die de binnenste haarcellen induceren.
Het afferente innervatiepatroon van het slakkenhuis. De illustratie wordt getoond door het vestibulaire (Reissner’s) membraan, naar “beneden” kijkend op het spiraalvormige orgaan (van Corti). De voornaamste vezelbundels zijn 1 en 2; 2a en 3a zijn interne spiraalvormige vezels; 4 zijn externe spiraalvormige vezels; 5 en 6 zijn radiale vezels die de binnenste haarcellen induceren.