Embryologie
Embryologicky se vestibulokochleární (akusticko-estibulární) ganglion zpočátku vyvíjí srostlé s ganglionem VII. nervu (obličejového). K oddělení těchto 3 složek dochází postupně. Neuroblasty vestibulárního a kochleárního (akustického) ganglia jsou odvozeny téměř výhradně z epitelu otocysty, na rozdíl od ostatních kraniálních senzorických ganglií, u nichž se na populaci neuroblastů podílejí jak gangliové, tak neurální hřebenové placky. Podpůrné Schwannovy a satelitní buňky jsou však stejně jako ve všech kraniálních gangliích výhradně původu z nervového hřebene a zřejmě pocházejí z ganglií lícního nervu (viz obrázek níže).
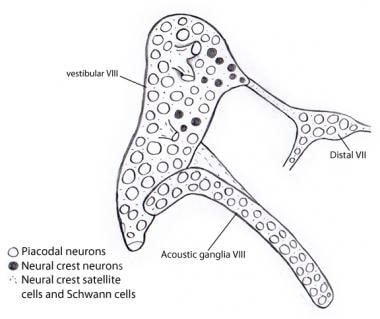 Ilustrace ukazující, že téměř všechny neurony ve vestibulárních a akustických gangliích pocházejí z otocysty jako placentární neurony. Naproti tomu všechny podpůrné a Schwannovy buňky jsou odvozeny z buněk nervového hřebene, pravděpodobně ze VII. nervového ganglia, ke kterému je vestibulokochleární ganglium původně připojeno.
Ilustrace ukazující, že téměř všechny neurony ve vestibulárních a akustických gangliích pocházejí z otocysty jako placentární neurony. Naproti tomu všechny podpůrné a Schwannovy buňky jsou odvozeny z buněk nervového hřebene, pravděpodobně ze VII. nervového ganglia, ke kterému je vestibulokochleární ganglium původně připojeno. Vzestupné (aferentní neboli projekční) dráhy sluchového nervu
Vzestupná dráha přenáší impulzy ze spirálního orgánu (Cortiho) do mozkové kůry (viz následující obrázek).
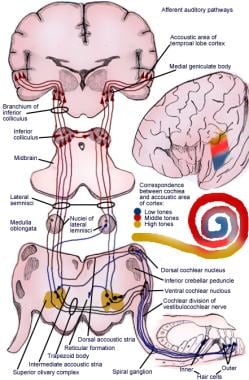 Ilustrace aferentní dráhy sluchového reflexu.
Ilustrace aferentní dráhy sluchového reflexu. Neurony prvního řádu sluchového systému
Vlákna kochleárního nervu vycházejí ze shluku nervových buněčných tělísek ve spirálním gangliu, které se nachází v modulu hlemýždě. Neurony spirálního ganglia jsou první ze 4 řádů neuronů mezi hlemýžděm a mozkem. Jsou to bipolární buňky, protože mají 2 sady výběžků neboli vláken, které vycházejí z opačných konců buněčných těl. Delší centrální vlákna, nazývaná také primární sluchová vlákna, tvoří kochleární nerv a kratší, periferní vlákna sahají k bázím vnitřních a vnějších vláskových buněk. Ze spirálního ganglia vycházejí radiálně do habenula perforata, což je řada malých otvorů pod vnitřními vláskovými buňkami. V tomto místě se demyelinizují a vstupují do spirálního orgánu (Cortiho).
Existuje pouze asi 30 000 těchto vláken a větší počet z nich – asi 95 % – inervuje vnitřní vláskové buňky. Zbytek prochází Cortiho tunelem a inervuje vnější vláskové buňky. Delší centrální výběžky bipolárních kochleárních neuronů se spojují a tvoří kochleární nervový kmen. Tato primární sluchová vlákna opouštějí modiolus přes vnitřní meatus a vstupují do prodloužené míchy. „Ústí“ vnitřního zvukovodu (IAC) se nazývá porus acusticus. Anatomické vztahy VII. a VIII. nervu v IAC a v oblasti mozečkového úhlu jsou důležitými anatomickými oblastmi souvisejícími s chirurgií baze lební a neurootology.
VII. a VIII. nerv jsou v celém svém intrakraniálním průběhu obaleny gliovou tkání. Tyto nervy jsou obklopeny Schwannovými buňkami začínajícími v IAC v blízkosti porus acusticus. Obersteinerova-Redlichova zóna je spojnicí glií a Schwannových nervů.
Scarpovo (vestibulární) ganglion leží přibližně uprostřed IAC. K rozdělení kraniálního nervu (CN) VIII na kochleární a vestibulární větev může dojít v mediálním segmentu IAC nebo v subarachnoidálním prostoru. Zadní polovina IAC je obsazena horní a dolní větví vestibulárního nervu. Kochleární nerv se nachází anteroinferiorně v kanálu. VII. nerv se nachází v anterosuperiorní části IAC. V horní části IAC odděluje obličejový a horní vestibulární nerv vertikální hřeben (Billův pruh). Příčný hřeben odděluje tuto horní část IAC od dolní části obsahující dolní vestibulární a kochleární oddíl.
Cerebelopontinní úhel je potenciální prostor v zadní jámě lební. Jeho hranice jsou následující:
-
Přední část: Zadní jamka spánkové kosti
-
Zadně: Přední plocha mozečku
-
Mediálně: Dolní oliva
-
Superiorálně: Dolní okraj ponsu a mozečkové stopky
-
Inferiorně: Mozečková mandle
Trojklanný nerv je viditelný výše než mozečkový úhel, zatímco IX., X. a XI. nerv probíhají níže. Mezi další důležité struktury v mozečkovém úhlu patří přední dolní mozečková tepna (AICA), flocculus a laterální otvor čtvrté komory (foramen of Luschka). Labyrintová tepna je obvykle větví AICA a zásobuje hlemýžď a labyrint.
Po vstupu do dřeně pokračují vlákna kochleárního nervu do kochleárního jádra. Kochleární jádro se skládá z 5 různých typů buněk, z nichž každý má odlišné morfologické a fyziologické vlastnosti, jako je odpověď na nástup podnětu, posun podnětu a frekvenční modulace. Kochleární jádro se dělí na dorzální a ventrální část. Kochleární vlákna se dělí na 2 hlavní svazky: Jedna skupina prochází laterálně a dorzálně k restiformnímu tělesu; druhá skupina zůstává mírně ventrálně a mediálně od restiformního tělesa a končí ve ventrálním kochleárním jádru. Bylo zjištěno, že vlákna vycházející z bazálních závitů hlemýždě končí v dorzální části dorzálního kochleárního jádra. Vlákna z apikálních částí hlemýždě končí ve ventrální části dorzálního kochleárního jádra a ve ventrálním jádře. Některá vlákna však procházejí k neuronům vyššího řádu dále po dráze, než dojde k jejich synapsi.
Neurony druhého, třetího a čtvrtého řádu sluchového systému
Buněčná těla neuronů druhého řádu leží v dorzálním a ventrálním kochleárním jádře. Některá vlákna z ventrálního kochleárního jádra procházejí přes střední čáru k buňkám horního olivového komplexu, zatímco jiná navazují spojení s olivovými buňkami téže strany. Horní olivový komplex je považován za první centrum ve vzestupném sluchovém systému, kde se sbíhají vstupy z obou uší. Společně tato vlákna tvoří lichoběžníkové těleso (kde se nacházejí neurony třetího řádu).
Vlákna z dorzálního kochleárního jádra překračují střední čáru a končí na buňkách jader laterálního lemnisku. Tam se k nim připojují vlákna z ventrálních kochleárních jader obou stran a z olivového komplexu. Sluchová jádra nad horním olivovým komplexem mohou být excitační nebo inhibiční se vstupy z každého ucha. Lemniscus je hlavní dráha, jejíž většina vláken končí v dolním koliculu, sluchovém centru středního mozku.
Dolní koliculus je komplexní jádro s nejméně 18 hlavními typy buněk a nejméně 5 oblastmi specializace. Podílí se na všech oblastech sluchového chování, včetně diferenciální citlivosti na frekvenci a intenzitu, hlasitost a binaurální slyšení. Ačkoli některá vlákna mohou colliculus obcházet a končit spolu s vlákny z colliculu na další vyšší úrovni, mediální genikulární těleso thalamu je místem, kde se nacházejí neurony čtvrtého řádu. Z mediálního genikulárního tělesa je uspořádaná projekce vláken do části kůry spánkového laloku.
Sluchový vstup
Primární sluchová kůra je první oblastí mozkové kůry, která přijímá sluchový vstup. U člověka a dalších primátů je primární akustickou oblastí v mozkové kůře horní příčná spánková gyri Heschlova, hřeben ve spánkovém laloku, na dolním okraji hluboké štěrbiny mezi spánkovým a temenním lalokem, známé jako laterální sulcus (Sylviova štěrbina).
Protože přibližně polovina vláken sluchových drah překračuje střední linii, zatímco ostatní vystupují na stejné straně mozku, je každé ucho zastoupeno v pravé i levé mozkové kůře. Z tohoto důvodu může být i při poškození sluchové kůry jedné strany úrazem nebo cévní mozkovou příhodou binaurální slyšení ovlivněno minimálně.
Vnímání zvuku je spojeno s pravým zadním horním spánkovým gyrem (STG). Horní spánkový gyrus obsahuje několik důležitých mozkových struktur, včetně Brodmannových 41 a 42, které označují umístění primární sluchové kůry, korové oblasti zodpovědné za vnímání základních charakteristik zvuku, jako je výška a rytmus.
Sluchová asociační oblast se nachází ve spánkovém laloku mozku, v oblasti zvané Wernickeho oblast nebo oblast 22. Tato oblast se nazývá „oblast 22“. Tato oblast v blízkosti laterálního mozkového sulku je důležitou oblastí pro zpracování akustických signálů tak, aby mohly být rozlišeny jako řeč, hudba nebo hluk.
Jak je běžné u talamokortikálních spojení, jádra v mediálním genikulárním tělese, která vysílají vlákna do sluchové kůry, přijímají také vlákna ze stejné oblasti kůry. Zhoršení sluchu v důsledku oboustranného poškození kůry zahrnujícího obě sluchové oblasti bylo zaznamenáno, ale je velmi vzácné. Bylo však prokázáno, že bilaterální léze spánkové mazové oblasti vyvolávají rozsáhlé účinky (korová hluchota, při níž je ovlivněno několik chování, včetně rozlišování řeči, lokalizace zvuku a detekce slabých, krátce trvajících signálů).
Následující obrázek znázorňuje vědomou a reflexní aferentní sluchovou dráhu.
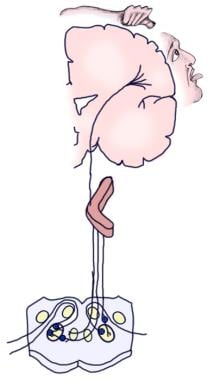 Ilustrace vědomé sluchové dráhy.
Ilustrace vědomé sluchové dráhy. 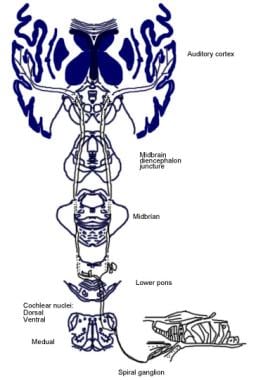 Ilustrace sluchové reflexní dráhy.
Ilustrace sluchové reflexní dráhy. Sestupné dráhy sluchového nervu
Kromě vědomé a reflexní aferentní sluchové dráhy existují také sestupné eferentní dráhy. Při vnímání zvuku hrají roli také eferentní projekce z mozku do hlemýždě. Obecně mají sestupné dráhy inhibiční účinek na vzestupná vlákna a mají tendenci zajišťovat autoregulaci sluchového systému. Každá reléová sluchová stanice je považována za duálně inervovanou, a poskytuje tak způsob, jak příchozí impulsy vnitřně ovlivnit, modifikovat nebo negovat.
Rovnoběžně s dráhou vzestupnou z kochleárních jader do kůry je dráha sestupná z kůry do kochleárních jader. V obou drahách zůstávají některá vlákna na stejné straně, zatímco jiná přecházejí přes střední čáru na opačnou stranu mozku. Existují také důkazy o „ostruhové“ linii stoupající z dorzálního kochleárního jádra do mozečku a další sestupující z dolního koliculu do mozečku.
Význam těchto mozkových spojení není jasný, ale mohou předcházet evolučnímu vývoji mozkové kůry. Z horního olivového komplexu, oblasti v prodloužené míše, vychází také dráha vláken zvaná olivokochleární svazek (viz obrázek níže). Tvoří eferentní systém neboli zpětnovazební smyčku, kterou se nervové impulzy, považované za inhibiční, dostávají k vláskovým buňkám.
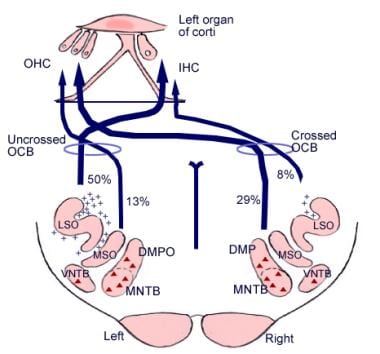 Ilustrace sestupných drah sluchového nervu. Křížky představují malé neurony olivokochleárního svazku (OCB); trojúhelníky představují velké neurony OCB. DMP, DMPO = dorzální mediální preolivární jádro; IHC = vnitřní vlásková buňka; LSO = laterální horní olivové jádro; MNTB = mediální jádro trapézového tělíska; MSO = mediální horní olivové jádro; OHC = vnější vlásková buňka; VNTB = ventrální jádro trapézového tělíska.
Ilustrace sestupných drah sluchového nervu. Křížky představují malé neurony olivokochleárního svazku (OCB); trojúhelníky představují velké neurony OCB. DMP, DMPO = dorzální mediální preolivární jádro; IHC = vnitřní vlásková buňka; LSO = laterální horní olivové jádro; MNTB = mediální jádro trapézového tělíska; MSO = mediální horní olivové jádro; OHC = vnější vlásková buňka; VNTB = ventrální jádro trapézového tělíska. Mikroskopická anatomie
Podrobnosti týkající se mikroskopické anatomie a inervace buněk ve spirálním orgánu (Cortiho) viz Vnitřní ucho . Následující obrázky zobrazují tuto oblast.
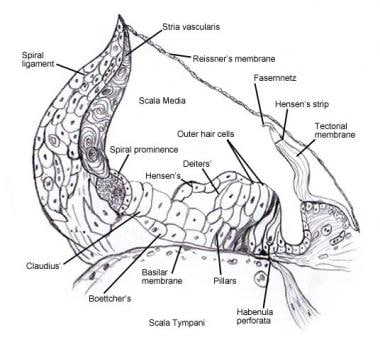 Spirální orgán (Cortiho).
Spirální orgán (Cortiho).  Skenovací elektronový mikrofotografický snímek horního povrchu Cortiho orgánu po odstranění tektoriální membrány. Jsou zde 3 řady vnějších vláskových buněk s charakteristickými do V uspořádanými stereociliemi. Zobrazena je také jedna řada vnitřních vláskových buněk s mírně zakřivenou řadou stereocilií.
Skenovací elektronový mikrofotografický snímek horního povrchu Cortiho orgánu po odstranění tektoriální membrány. Jsou zde 3 řady vnějších vláskových buněk s charakteristickými do V uspořádanými stereociliemi. Zobrazena je také jedna řada vnitřních vláskových buněk s mírně zakřivenou řadou stereocilií. 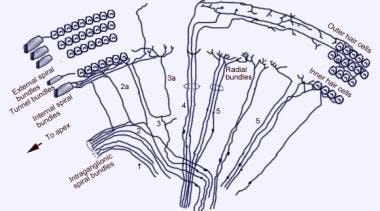 Schéma aferentní inervace hlemýždě. Obrázek je zobrazen přes vestibulární (Reissnerovu) membránu při pohledu „dolů“ na spirální orgán (Cortiho). Hlavní svazky vláken jsou 1 a 2; 2a a 3a jsou vnitřní spirální vlákna; 4 jsou vnější spirální vlákna; 5 a 6 jsou radiální vlákna inervující vnitřní vláskové buňky.
Schéma aferentní inervace hlemýždě. Obrázek je zobrazen přes vestibulární (Reissnerovu) membránu při pohledu „dolů“ na spirální orgán (Cortiho). Hlavní svazky vláken jsou 1 a 2; 2a a 3a jsou vnitřní spirální vlákna; 4 jsou vnější spirální vlákna; 5 a 6 jsou radiální vlákna inervující vnitřní vláskové buňky.