Embriologie
Embriologic, ganglionul vestibulococlear (acusticovestibular) se dezvoltă inițial fuzionat cu ganglionul nervului al VII-lea (facial). Separarea celor 3 componente are loc treptat. Neuroblastele ganglionilor vestibulari și cohleari (acustici) derivă aproape exclusiv din epiteliul otocistului, spre deosebire de alți ganglioni senzoriali cranieni în care atât placodele ganglionare, cât și cele ale crestei neuronale contribuie în mare măsură la populațiile de neuroblaste. Cu toate acestea, celulele Schwann și celulele satelit de susținere, ca în toate ganglionii cranieni, sunt în întregime de origine din creasta neurală, aparent provenind din ganglionul nervului facial (a se vedea imaginea de mai jos).
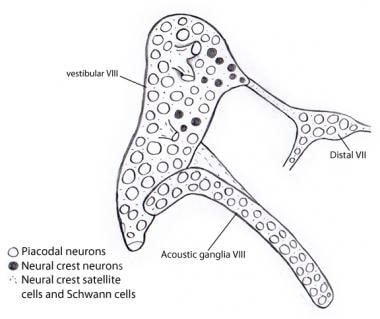 Ilustrație care arată că aproape toți neuronii din ganglionii vestibulari și acustici sunt derivați din otocist ca neuroni placodali. În schimb, toate celulele de susținere și celulele Schwann derivă din celulele crestei neurale, posibil din ganglionul al VII-lea nervos la care este atașat inițial ganglionul vestibulococlear.
Ilustrație care arată că aproape toți neuronii din ganglionii vestibulari și acustici sunt derivați din otocist ca neuroni placodali. În schimb, toate celulele de susținere și celulele Schwann derivă din celulele crestei neurale, posibil din ganglionul al VII-lea nervos la care este atașat inițial ganglionul vestibulococlear. Calea ascendentă (aferentă sau proiectivă) a nervului auditiv
Calea ascendentă transmite impulsuri de la organul spiralat (de Corti) la cortexul cerebral (vezi imaginea următoare).
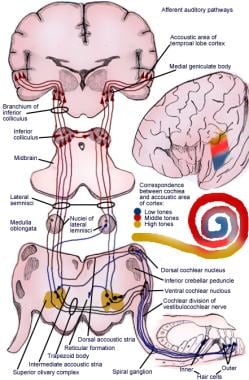 Ilustrație a căii aferentă a reflexului auditiv.
Ilustrație a căii aferentă a reflexului auditiv. Neuronii de prim ordin ai sistemului auditiv
Fibrele nervului cohlear își au originea într-o agregare de corpuri celulare nervoase din ganglionul spiralat, situat în modiolul cohleei. Neuronii din ganglionul spiral sunt primii neuroni de 4 ordine între cohlee și creier. Aceștia sunt celule bipolare, deoarece au 2 seturi de procese, sau fibre, care se extind de la capetele opuse ale corpurilor celulare. Fibrele centrale mai lungi, numite și fibre auditive primare, formează nervul cohlear, iar fibrele periferice, mai scurte, se extind până la bazele celulelor ciliate interne și externe. Acestea se extind radial de la ganglionul spiral până la habenula perforata, o serie de găuri mici sub celulele ciliate interne. În acest punct, ele devin demielinizate pentru a intra în organul spiralat (de Corti).
Există doar aproximativ 30.000 dintre aceste fibre, iar cel mai mare număr dintre ele – aproximativ 95% – inervează celulele ciliate interne. Restul traversează tunelul lui Corti pentru a inerva celulele ciliate externe. Procesele centrale mai lungi ale neuronilor cohleari bipolari se unesc pentru a forma trunchiul nervului cohlear. Aceste fibre auditive primare ies din modiolus prin meatul intern și intră în medulla oblongata. „Gura” canalului auditiv intern (CAI) se numește porus acusticus. Relația anatomică a nervilor VII și VIII în CAI și în regiunea unghiului cerebellopontin sunt zone anatomice importante legate de chirurgia bazei craniului și de neuro-otologi.
Nervii VII și VIII sunt înveliți în țesut glial pe tot parcursul traseului lor intracranian. Acești nervi sunt înconjurați de celule Schwann începând în IAC, aproape de porus acusticus. Zona Obersteiner-Redlich este joncțiunea glială-Schwann.
Ganglionul Scarpa (vestibular) se află aproximativ în mijlocul IAC. Diviziunea nervului cranian (CN VIII) în ramurile cohleară și vestibulară poate avea loc în segmentul medial al CAI sau în spațiul subarahnoidian. Jumătatea posterioară a CAI este ocupată de ramurile superioare și inferioare ale nervului vestibular. Nervul cohlear este situat antero-inferior în canal. Al VII-lea nerv este situat în porțiunea antero-superioară a CAI. O creastă verticală (Bill bar) separă nervii vestibulari facial și superior în partea superioară a CAI. Creasta transversală separă această porțiune superioară a CAI de partea inferioară care conține diviziunile vestibulare inferioare și cohleare.
Unghiul cerebellopontin este un spațiu potențial în fosa craniană posterioară. Limitele sale sunt următoarele:
-
Anterior: Fosa posterioară a osului temporal
-
Posterior: Suprafața anterioară a cerebelului
-
Medial: Oliveta inferioară
-
Superior: Marginea inferioară a ponsului și a pedunculului cerebelos
-
Inferior: Amigdala cerebelară
Nervul trigemen este vizibil superior unghiului cerebelopontin, în timp ce nervii IX, X și XI se desfășoară inferior. Alte structuri importante din cadrul unghiului cerebellopontin includ artera cerebelară inferioară anterioară (AICA), floculul și deschiderea laterală a celui de-al patrulea ventricul (foramenul lui Luschka). Artera labirintică este de obicei o ramură a AICA și alimentează cohleea și labirintul.
După ce intră în măduvă, fibrele nervoase cohleare se îndreaptă spre nucleul cohlear. Nucleul cohlear este format din 5 tipuri de celule distincte, fiecare cu caracteristici morfologice și fiziologice distincte, cum ar fi răspunsul la debutul stimulului, la decalajul stimulului și la modularea frecvenței. Nucleul cohlear este împărțit în părțile dorsală și ventrală. Fibrele cohleare se împart în 2 fascicule principale: Un grup trece lateral și dorsal spre corpul restiform; celălalt grup rămâne ușor ventral și medial față de corpul restiform și se termină în nucleul cohlear ventral. S-a constatat că fibrele care provin din spirele bazale ale cohleei se termină în în partea dorsală a nucleului cohlear dorsal. Fibrele care provin din părțile apicale ale cohleei se termină în partea ventrală a nucleului cohlear dorsal și în nucleul ventral. Cu toate acestea, unele fibre trec la neuronii de ordin superior mai departe de-a lungul căii înainte de a face sinapsă.
Neuronii de ordinul doi, trei și patru ai sistemului auditiv
Corpurile celulare ale neuronilor de ordinul doi se află în nucleii cohleari dorsali și ventrali. Unele fibre din nucleul cohlear ventral trec peste linia mediană către celulele complexului olivionar superior, în timp ce altele fac legătura cu celulele olivare din aceeași parte. Complexul olivionar superior este considerat primul centru din sistemul auditiv ascendent, unde converg intrările de la ambele urechi. Împreună, aceste fibre formează corpul trapezoidal (unde sunt localizați neuronii de ordinul al treilea).
Fibrele din nucleul cohlear dorsal traversează linia mediană pentru a se termina pe celulele nucleilor din lemniscusul lateral. Acolo li se alătură fibrele de la nucleii cohleari ventrali de ambele părți și de la complexul olivar. Nucleii auditivi de deasupra complexului olivionar superior pot fi excitatori sau inhibitori, cu intrări de la fiecare ureche. Lemniscul este un tract major, ale cărui fibre se termină în cea mai mare parte în coliculul inferior, centrul auditiv al mezencefalului.
Coliculul inferior este un nucleu complex cu cel puțin 18 tipuri de celule majore și cel puțin 5 zone de specializare. Este implicat în toate domeniile comportamentului auditiv, inclusiv în sensibilitatea diferențială pentru frecvență și intensitate, intensitate sonoră și auz binaural. Deși unele fibre pot ocoli coliculul și se termină, împreună cu fibrele din colicul, la nivelul imediat superior, corpul geniculat medial al talamusului este locul unde sunt localizați neuronii de ordinul al patrulea. Din corpul geniculat medial, există o proiecție ordonată a fibrelor către o porțiune din cortexul lobului temporal.
Intrare auditivă
Cortexul auditiv primar este prima regiune a cortexului cerebral care primește intrare auditivă. La om și la alte primate, zona acustică primară din cortexul cerebral este girusul temporal transversal superior al lui Heschl, o creastă din lobul temporal, pe buza inferioară a despicăturii profunde dintre lobii temporal și parietal, cunoscută sub numele de sulcus lateral (fisura Sylviană).
Pentru că aproximativ jumătate din fibrele căilor auditive traversează linia mediană, în timp ce altele urcă pe aceeași parte a creierului, fiecare ureche este reprezentată atât în cortexul drept, cât și în cel stâng. Din acest motiv, chiar și atunci când zona corticală auditivă a unei părți este lezată prin traumatism sau accident vascular cerebral, auzul binaural poate fi afectat în mod minim.
Percepția sunetului este asociată cu girusul temporal superior posterior drept (STG). Girusul temporal superior conține mai multe structuri importante ale creierului, inclusiv Brodmann 41 și 42, care marchează localizarea cortexului auditiv primar, regiunea corticală responsabilă de senzația caracteristicilor de bază ale sunetului, cum ar fi înălțimea și ritmul.
Azona de asociere auditivă este localizată în cadrul lobului temporal al creierului, într-o zonă numită zona Wernicke sau zona 22. Această zonă, în apropierea sulcusului cerebral lateral, este o regiune importantă pentru procesarea semnalelor acustice, astfel încât acestea să poată fi distinse ca vorbire, muzică sau zgomot.
Cum este obișnuit pentru conexiunile talamocorticale, nucleii din cadrul corpului geniculat medial care trimit fibre către cortexul auditiv primesc, de asemenea, fibre din aceeași zonă a cortexului. A fost raportată o deficiență de auz datorată unei leziuni corticale bilaterale care implică ambele zone auditive, dar este extrem de rară. Cu toate acestea, s-a demonstrat că leziunile bilaterale ale limbii temporale produc efecte de amploare (surditate corticală, în care sunt afectate mai multe comportamente, inclusiv discriminarea vorbirii, localizarea sunetului și detectarea semnalelor slabe, de scurtă durată).
În următoarea imagine sunt reprezentate căile auditive aferente conștiente și reflexe.
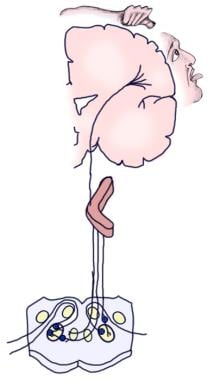 Ilustrație a căii auditive conștiente.
Ilustrație a căii auditive conștiente. 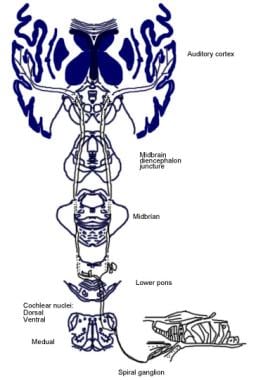 Ilustrație a căii auditive reflexe.
Ilustrație a căii auditive reflexe. Căile descendente ale nervului auditiv
În afară de căile auditive aferente conștiente și reflexe, există și căi eferente descendente. Proiecțiile eferente de la creier către cohlee joacă, de asemenea, un rol în percepția sunetului. În general, căile descendente au un efect inhibitor asupra fibrelor ascendente și au tendința de a asigura autoreglarea sistemului auditiv. Se consideră că fiecare stație auditivă de releu este inervată dual, oferind astfel o modalitate prin care impulsurile primite pot fi influențate, modificate sau negate pe plan intern.
În paralel cu calea care urcă de la nucleii cohleari la cortex există o cale care coboară de la cortex la nucleii cohleari. În ambele căi, unele dintre fibre rămân pe aceeași parte, în timp ce altele traversează linia mediană spre partea opusă a creierului. Există, de asemenea, dovezi ale existenței unei linii de „pinteni” care urcă de la nucleul cohlear dorsal la cerebel și o alta care coboară de la coliculul inferior la cerebel.
Semnificația acestor conexiuni cerebrale nu este clară, dar este posibil ca ele să fie anterioare dezvoltării evolutive a cortexului cerebral. Din complexul olivar superior, o regiune din bulbul rahidian, provine, de asemenea, un tract de fibre numit fasciculul olivococlear (vezi imaginea de mai jos). Acesta constituie un sistem eferent, sau o buclă de feedback, prin care impulsurile nervoase, despre care se crede că sunt inhibitorii, ajung la celulele ciliate.
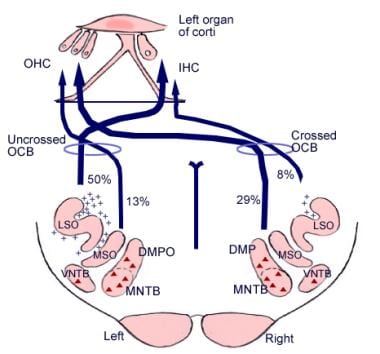 Ilustrație a căilor descendente ale nervului auditiv. Crucile reprezintă neuronii mici din fasciculul olivocochlear (OCB); triunghiurile reprezintă neuronii mari din OCB. DMP, DMPO = nucleu preolivar medial dorsal; IHC = celulă ciliată internă; LSO = nucleu olivionar superior lateral; MNTB = nucleu medial al corpului trapezoidal; MSO = nucleu olivionar superior medial; OHC = celulă ciliată externă; VNTB = nucleu ventral al corpului trapezoidal.
Ilustrație a căilor descendente ale nervului auditiv. Crucile reprezintă neuronii mici din fasciculul olivocochlear (OCB); triunghiurile reprezintă neuronii mari din OCB. DMP, DMPO = nucleu preolivar medial dorsal; IHC = celulă ciliată internă; LSO = nucleu olivionar superior lateral; MNTB = nucleu medial al corpului trapezoidal; MSO = nucleu olivionar superior medial; OHC = celulă ciliată externă; VNTB = nucleu ventral al corpului trapezoidal. Anatomie microscopică
Vezi Urechea internă pentru detalii privind anatomia microscopică și inervația celulelor din organul spiralat (de Corti) . Următoarele imagini înfățișează această zonă.
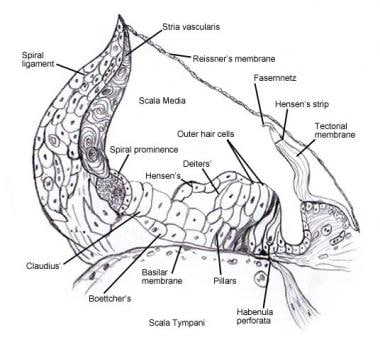 Organul spiralat (de Corti).
Organul spiralat (de Corti).  Micrografie electronică de scanare a suprafeței superioare a organului lui Corti după îndepărtarea membranei tectoriale. Există 3 rânduri de celule ciliate externe cu stereociile lor caracteristice dispuse în V. Se arată, de asemenea, un singur rând de celule ciliate interne cu un rând ușor curbat de stereocili.
Micrografie electronică de scanare a suprafeței superioare a organului lui Corti după îndepărtarea membranei tectoriale. Există 3 rânduri de celule ciliate externe cu stereociile lor caracteristice dispuse în V. Se arată, de asemenea, un singur rând de celule ciliate interne cu un rând ușor curbat de stereocili. 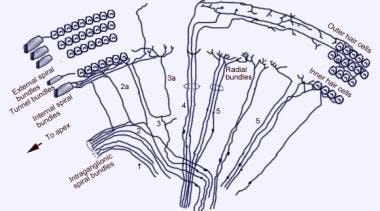 Schema de inervație aferentă a cohleei. Ilustrația este prezentată prin membrana vestibulară (Reissner), privind „în jos” spre organul spiralat (de Corti). Principalele fascicule de fibre sunt 1 și 2; 2a și 3a sunt fibre spiralate interne; 4 sunt fibre spiralate externe; 5 și 6 sunt fibre radiale care inervează celulele ciliate interne.
Schema de inervație aferentă a cohleei. Ilustrația este prezentată prin membrana vestibulară (Reissner), privind „în jos” spre organul spiralat (de Corti). Principalele fascicule de fibre sunt 1 și 2; 2a și 3a sunt fibre spiralate interne; 4 sunt fibre spiralate externe; 5 și 6 sunt fibre radiale care inervează celulele ciliate interne.