Introduktion
Den menneskelige hjerne gennemgår en omfattende modning gennem hele livet for at fremme den kognitive udvikling. Myeliniseringen af axoner i hele nervesystemet er en af disse afgørende modningsprocesser. I centralnervesystemet (CNS) strækker gliaceller kaldet oligodendrocytter mange processer ud i deres omgivende miljø, som koncentreret vikler membranen omkring axoner for at danne myelinskeder. Myelinskederne muliggør den hurtige saltatoriske ledning af aktionspotentialer ved at lokalisere spændingsafhængige Na+-kanaler til korte huller mellem tilstødende skeder (kendt som Ranvier-knuderne) og ved at fungere som elektriske isolatorer. Axoner, der er fuldt myeliniserede i hele deres længde, leder impulser mange gange hurtigere end ikke-myeliniserede axoner med samme tværsnitsstørrelse (Waxman, 1980). Derfor leder myeliniserede neurale kredsløb information meget hurtigere end ikke-myeliniserede kredsløb. Mennesker fødes med et stort set umyeliniseret CNS, og oligodendrocytpopulationen udvider sig dramatisk efter fødslen med udbredt myelinisering i de første par år af barndommen. Myeliniseringen fortsætter gennem ungdomsårene og ind i voksenalderen på en karakteristisk spatiotemporal måde, der korrelerer med fremkomsten og opretholdelsen af en korrekt kredsløbsfunktion. F.eks. er modningen af hvidt stof (de myelinrige områder af CNS) samtidig med udviklingen af kognitive processer i barndommen, såsom informationsbehandlingshastighed (Mabbott et al., 2006; Scantlebury et al., 2014). Desuden ses myelinpatologi/abnormaliteter ikke kun i den demyeliniserende sygdom multipel sklerose, men også i flere neurodegenerative sygdomme (Kang et al., 2013; Huang et al., 2015) og neuroudviklingsforstyrrelser (Takahashi et al., 2011). Myelinisering af individuelle axoner er imidlertid ikke et “alt-eller-intet”-fænomen. Axoner i CNS udviser omfattende variation i antallet af myelinskeder, skedernes længde, skedernes tykkelse og fordelingen langs deres længde. Der findes mange forskellige mønstre af myelinisering; for eksempel er axoner med sparsomt myeliniserede regioner blevet beskrevet i juvenil og voksen musekortex (Tomassy et al., 2014; Hill et al., 2018; Hughes et al., 2018). Ændring af nogen af disse kappeparametre har forudsigelige virkninger på ledningshastigheden af det underliggende axon – derfor kan det være særligt vigtigt at etablere et specifikt mønster af myelinisering langs et axon for at finjustere kredsløbsfunktionen. For eksempel udviser axoner i den auditive hjernestamme hos gerbiler progressivt kortere myelinskeder langs distale regioner for at sikre den præcise timing af signalets ankomst for at lette lydlokalisering (Ford et al., 2015). Subtile ændringer i det overordnede mønster af myelinisering langs et axon (enten via tilføjelse af nyt myelin eller remodellering af eksisterende myelin) kunne ændre timingen af neurale impulser i kredsløb i dyb grad. Hvis myelin er tilpasningsdygtigt, kan ændring af sådanne myeliniseringsmønstre være en effektiv mekanisme til regulering af kredsløbsfunktionen gennem hele livet.
Nylige beviser tyder på, at myelin kan være tilpasningsdygtigt som reaktion på kredsløbsaktivitet. Diffusionstensorafbildning af hele hjernen kan bruges til at måle brede ændringer i den myelinrige hvide substans over tid – sådanne eksperimenter hos mennesker og gnavere har vist, at indlæring af en ny opgave korrelerer med ændringer i den hvide substans i relevante hjerneområder (Scholz et al., 2009; Sampaio-Baptista et al., 2013). Analyser på celleniveau i dyremodeller viser, at produktionen af nye myeliniserende oligodendrocytter er nødvendig for effektiv motorisk indlæring (McKenzie et al., 2014; Xiao et al., 2016). Det er i øjeblikket en hypotese, at aktivitet i neurale kredsløb kan udløse ændringer i myelin; en omfattende forskning har vist, at neuronal aktivitet kan påvirke proliferationen af oligodendrocytprækursorceller (OPC’er), differentieringen af oligodendrocytter og dannelsen og væksten af myelinskeder. Denne forskning, herunder beviser for de involverede molekylære signaler, er blevet grundigt gennemgået andre steder (Fields, 2015; Almeida og Lyons, 2017; Mount og Monje, 2017). Neuronal aktivitet kunne drive ændringer i myelin, som igen kunne ændre ledningshastighederne for at finjustere de tidspunkter, der ligger til grund for kredsløbsfunktionen.
Derimod ved vi stadig ikke, om eller hvordan myeliniseringen af kredsløb reguleres dynamisk gennem hele livet. Gnaverarbejde viser, at der dannes nye oligodendrocytter i hele CNS selv i voksenalderen (Young et al., 2013), og OPC’er findes i den menneskelige voksne hjerne (Chang et al., 2000). Kulstofdateringsanalyser af menneskeligt væv har identificeret voksenfødte oligodendrocytter i cortexen, selv om de samme analyser viste, at størstedelen af oligodendrocytterne i corpus callosum stammer fra den tidlige barndom (Yeung et al., 2014). Imidlertid tyder neuroimaging-undersøgelser hos mennesker, der korrelerer strukturelle ændringer i den hvide substans med opgavelæring, på, at der kan dannes nyt myelin hele livet igennem. En så langvarig myelinisering ville i princippet kræve livslang produktion af oligodendrocytter, da individuelle myeliniserende oligodendrocytter har et begrænset tidsvindue på kun få timer til at indlede dannelsen af nye kapper (Watkins et al., 2008; Czopka et al., 2013), og kappeantallet pr. oligodendrocyt synes stabilt over tid (Tripathi et al., 2017). Et forbehold bemærket af Mount og Monje (2017) er, at “fødselsdatoen” i kulstof-dateringsforsøget (som identificerer tidspunktet for DNA-replikation under celledeling), afspejler OPC’en, ikke nødvendigvis den differentierede oligodendrocyt. Dette er vigtigt i betragtning af beviser for, at OPC’er kan differentiere direkte til oligodendrocytter uden celledeling, i det mindste hos gnavere (Hughes et al., 2013). OPC’er i corpus callosum kunne differentiere direkte til oligodendrocytter mange år efter deres terminale celledeling; derfor kan tidspunktet for differentiering af disse nye oligodendrocytter ikke bestemmes ved hjælp af kulstofdatering, og derfor kan Yeung et al. (2014) have undervurderet hastigheden af oligodendrocytproduktion i den voksne menneskelige hjerne. Vi har stadig meget at lære om de relative bidrag fra oligodendrocytgenerering og myelinremodellering til CNS udvikling gennem hele livet.
For fuldt ud at forstå den præcise dynamik af oligodendrogenese, myelindannelse og myelinremodellering gennem forskellige stadier af livet repræsenterer longitudinal billeddannelse med høj opløsning den gyldne standardtilgang. Her giver vi en oversigt over nylige in vivo billeddannelsesundersøgelser, der begynder at klarlægge myeliniseringens dynamik, hvilket også vil give os mulighed for at begynde at forstå, hvordan en sådan dynamik kan påvirke neurale kredsløbsfunktioner.
De novo Myelinisering
For at begynde at få endeligt svar på, hvordan oligodendrocytter dannes, og hvordan myelin dannes og dynamisk remodelleres in vivo, har to nylige undersøgelser anvendt gentagen to-foton billeddannelse af den somatosensoriske cortex hos mus over længere tidsperioder. Hughes et al. (2018) afbildede cortexen af transgene reportermus med fluorescerende mærkede oligodendrogliale lineageceller fra tidlig voksenalder, gennem middelalder og alderdom (ca. P720). De fandt, at oligodendrocytpopulationen fortsætter med at ekspandere, og at den kortikale oligodendrocyttæthed næsten fordobles mellem unge voksne og midaldrende stadier (Figur 1A). Dette blev ledsaget af en over dobbelt så stor stigning i antallet af kortikale myelinskeder. Men hvordan øges antallet af oligodendrocytter? I den tidlige postnatale udvikling produceres der mange oligodendrocytter, men kun en delmængde overlever og fortsætter med at myelinisere axoner (Barres et al., 1992). Dette synes at være tilsvarende i voksenalderen – ved at følge individuelle kortikale OPC’er i den voksne cortex i op til 50 dage viste Hughes et al. (2018), at størstedelen af de nyligt differentierede oligodendrocytter gennemgår celledød, hvor kun 22% overlever og forpligter sig til myelinisering (Figur 1B). Det er fortsat ukendt, hvilken andel af de nydifferentierede oligodendrocytter der genereres efter OPC-division versus direkte differentiering. Men når oligodendrocytterne først er gået i gang med myelinisering, forbliver de stabile, og der er ingen tegn på, at myeliniserende oligodendrocytter undergår celledød i løbet af en 50-dages billeddannelsesperiode.
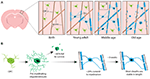
FIGUR 1. Oligodendrocyt- og myelin-dynamik i pattedyrs cortex gennem hele livet. (A) Oligodendrocytprækursorceller (OPC’er) genererer kontinuerligt nye myeliniserende oligodendrocytter (OL’er) i den somatosensoriske cortex fra fødslen og frem til den midaldrende alder. OL-populationen falder derefter i alderdommen, hvilket ledsages af en reduktion af myelin-dækningen. (B) Lineage-tracing af enkelte OPC’er viser, at selv om der kontinuerligt produceres præmyeliniserende OL’er i voksenalderen, er det kun ca. 20 %, der overlever til at myelinisere. De fleste myelinskeder, når de først er dannet, er stabile i længden over længere tid, hvilket indikerer, at der normalt er meget lidt remodellering af det eksisterende myelin. Sammenfatning af data fra Hill et al. (2018) og Hughes et al. (2018).
Sådan brugte Hill et al. (2018) transgene reportere af oligodendrocytter og den mærkningsfrie spektrale konfokale refleksionsteknik (SCoRe) til at afbilde myelin langs axoner (Schain et al., 2014) i den somatosensoriske cortex hos juvenile, unge voksne, midaldrende og gamle mus (P950). De fandt også, at antallet af oligodendrocytter fortsætter med at vokse i voksenalderen op til P650, og at oligodendrocytterne er stabile i middelalderen i op til 80 dage efter billeddannelse. De fandt, at myeliniseringen af cortex også topper i middelalderen ved P650, og at oligodendrocyttætheden falder betydeligt fra sit højdepunkt (ved P650) til meget høj alder (P950) (figur 1A). Dette blev afspejlet i en reduktion af myelin-dækningen af lag I kortikale axoner mellem P650 og P950. Langtidsoverlevelsen af oligodendrocytter kan variere mellem forskellige dele af CNS. Tripathi et al. (2017) mærkede myeliniserende oligodendrocytter ved P60 i mus og talte derefter, hvor mange mærkede celler der overlevede indtil P605 i flere CNS-regioner. De fandt, at i rygmarven og den motoriske cortex overlevede 60-70 % af de P60-mærkede celler, mens over 90 % af de P60-mærkede celler overlevede i corpus callosum. Reduktionen i antallet af oligodendrocytter og myelinisering i visse CNS-regioner med alderen rejser spændende spørgsmål om den rolle, som myelintab spiller i aldersrelateret kognitiv svækkelse. MRI-analyse viser, at mikrostruktur af hvid substans korrelerer med flydende intelligens (Ritchie et al., 2015), men også at denne mikrostruktur af hvid substans forringes med stigende alder (Cox et al., 2016). Efterfølgende aldersrelateret myelin tab kan føre til nedsat kognitiv funktion på grund af dysregulering af myeliniserede kredsløb.
Kunne genereringen af nye oligodendrocytter (og efterfølgende nyt myelin) i den voksne cortex være lydhør over for kredsløbsaktivitet? Tidligere forskning har vist, at reduktion af sensorisk input ved at fjerne knurhår fra mus fører til reduceret oligodendrogenese i den somatosensoriske cortex (Hill et al., 2014). For at undersøge dette yderligere, gav Hughes et al. (2018) voksne (P365) mus med sensorisk stimulering i 3 uger ved at hænge perler i dyreburene for gentagne gange at stimulere deres whiskers og dermed den somatosensoriske cortex. Ved at afbilde den somatosensoriske cortex før og efter de 3 uger viste de, at sensorisk stimulering øger antallet af oligodendrocytter, potentielt på grund af den øgede overlevelse af nydifferentierede celler. Kougioumtzidou et al. (2017) gav yderligere beviser for, at kredsløbsaktivitet kan være vigtig i reguleringen af celleoverlevelse – de viste, at tab af AMPA-receptorunderenheder 2, 3 og 4 i OPC’er fører til reduceret overlevelse af oligodendrocytter. Dette tyder på, at de novo myelinisering kan moduleres af kortikal kredsløbsaktivitet gennem hele livet, måske for at finjustere funktionen af de samme kredsløb.
Mange spørgsmål mangler stadig at blive besvaret: Hvad er effekten af oligodendrogenese og ny myelinisering på den faktiske kredsløbsfunktion? Forbedrer neuronal aktivitet den langsigtede overlevelse af myeliniserende oligodendrocytter på lang sigt? Det er muligt, at tabet af oligodendrocytter i alderdommen skyldes aldersrelateret reduktion i neuronal aktivitet, hvilket igen kan påvirke den samlede oligodendrocytoverlevelse. Alternativt kan det være, at oligodendrocytter har en begrænset levetid uafhængigt af neuronal aktivitet (enten iboende programmeret eller påvirket af andre extrinsiske signaler, der er forbundet med aldring). I begge tilfælde kan kredsløbsstimulering bidrage til at afhjælpe aldersrelateret myelintab ved enten at fremme overlevelsen af eksisterende oligodendrocytter eller stimulere produktionen af nye oligodendrocytter. Dette kunne igen have betydelige implikationer i behandlingen og forebyggelsen af aldersassocieret kognitiv svækkelse.
Aktivitetsmedieret oligodendrogenese er ikke begrænset til den somatosensoriske cortex – unge voksne mus, der gennemgår motorisk indlæring, viser også en stigning i antallet af nydifferentierede oligodendrocytter i den motoriske cortex (Xiao et al., 2016). Hvad med andre områder af CNS? Mange kortikale axoner projicerer via corpus callosum, og derfor kan stimulering af kortikale kredsløb signalere til både kortikale og callosale OPC’er. To gnaverundersøgelser har vist, at stimulering af kortikale neuroner inducerer oligodendrogenese i corpus callosum. Gibson et al. (2014) optogenetisk stimulerede lag V-projektionsneuroner i den præmotoriske cortex og fandt en stigning i OPC-proliferation i både den præmotoriske cortex og corpus callosum. Dette førte til en stigning i antallet af oligodendrocytter og skindtykkelse 4 uger efter stimulering. For nylig brugte Mitew et al. (2018) Designer Receptors Exclusively Activated by Designer Drugs til at stimulere lag 2/3 somatosensoriske neuroner og observerede også øget OPC-proliferation, oligodendrogenese og tykkere myelinskeder i corpus callosum hos både juvenile og voksne mus. De påviste også, at nye oligodendrocytter fortrinsvis danner myelinskeder på de aktive axoner. Dette tyder på, at aktivitetsinduceret de novo myelinisering i princippet kan være rettet mod aktive axoner/kredsløb. Det er fortsat uvist, hvor langvarige ændringer af myelin som følge af neuronal aktivitet kan være. Den langsigtede overlevelse af myeliniserende celler, som Tripathi et al. (2017), Hill et al. (2018) og Hughes et al. (2018) har bemærket, tyder på, at når først en oligodendrocyt danner myelinskeder, er det sandsynligt, at den vil overleve, selv om neuronale aktivitetsniveauer vender tilbage til baseline. Hvorvidt myelinskederne selv ændrer sig, når den neuronale aktivitet vender tilbage til normale niveauer, kræver mere undersøgelse af individuelle skinddynamikker, hvilket diskuteres nedenfor.
Det er således muligt, at livslang de novo myelinisering kan forekomme i mange CNS-regioner, hvor der er tilstrækkeligt med umyeliniseret plads i de axoner, der er egnede til myelinisering. Det er imidlertid fortsat uklart, i hvilket omfang oligodendrogenesen fortsætter i forskellige områder af den voksne menneskelige hjerne. Kulstofdateringsanalyse tyder på, at de fleste oligodendrocytter i corpus callosum-trakten dannes i den tidlige barndom (Yeung et al., 2014). Immunohistokemisk analyse af humant hjernevæv ved hjælp af en ny markør for nydifferentierede oligodendrocytter (BCAS1) viser nye oligodendrocytter i den frontale cortex selv efter middellevealderen, men meget få nye oligodendrocytter i den hvide substans efter det tredje årti af livet (Fard et al., 2017). Denne forskel i oligodendrogenese mellem arter kan være et resultat af skalaen. Hughes et al.’s (2013) data fra gnavere tyder på, at oligodendrocytter dannes i et enormt overskud med kontinuerlig beskæring af næsten 80 % af cellerne. I betragtning af energiomkostningerne ved en sådan proces, er denne mekanisme bæredygtig gennem hele livet i et organ af størrelsen af den menneskelige hjerne? Måske er der i den menneskelige hjerne begrænset overproduktion af oligodendrocytter, fordi der er behov for en mere langvarig myelinisering af det større CNS, eller fordi signaler som neuronal aktivitet stimulerer OPC’er til at differentiere sig til oligodendrocytter efter behov.
Myelinomdannelse
Omdannelsen af eksisterende myelinskeder kunne ændre ledningsegenskaberne uden at der er behov for at generere nye oligodendrocytter eller myelin. Ændring af længden af eksisterende myelinskeder kunne ændre myelinets dækning langs et axon og afstanden mellem Ranvier-knuderne (hvilket begge ville påvirke ledningshastighederne). Desuden kan selv en meget subtil myelinomdannelse ændre længden af selve knuderne. Det er for nylig blevet vist, at knudelængden kan variere betydeligt i synsnerven og i cortex, og at ændring af knudelængderne langs et axon i princippet også kan ændre ledningshastigheden betydeligt (Arancibia-Cárcamo et al., 2017). Hvorvidt ændringer i node of Ranvier primært er drevet af myelinisering eller reorganisering af selve axonet mangler stadig at blive bestemt.
Både Hill et al. (2018) og Hughes et al. (2018) udførte longitudinelle undersøgelser af individuelle myelinskeder i musens somatosensoriske cortex i flere uger for at vurdere, om skede længder er dynamisk reguleret. Hill et al. (2018) fandt, at i det tidlige voksenliv (P90-120), selv om nogle kapper udviser udvidelse eller krympning, var 81% af de observerede kapper stabile. Flere kapper kan blive stabile i længde med alderen; Hughes et al. (2018) fulgte kapper i ældre (P365) dyr og så, at 99% af kapperne forblev stabile over 3 uger (Figur 1B).
Sammenlignende kappelængde stabilitet er også blevet beskrevet andre steder; Auer et al. (2018) brugte larve zebrafisk til at undersøge, om individuelle kapper kan ændre længde over tid ved at udføre tidsforløb live billeddannelse af fluorescensmærkede myelinskeder. De fandt, at individuelle kapper gennemgår hurtig, men variabel vækst i de første par dage efter dannelsen, før de stabiliserer deres kappelængder. Når de først er stabiliseret, fortsætter kapperne kun med at vokse i takt med dyrets samlede vækst.
Hvorfor ændrer nogle kapper i cortex sig i længde, mens andre ikke gør det? Dette kan afspejle forskellighed i kravene fra forskellige neurale kredsløb. Axonal diversitet er blevet observeret under den indledende myelinisering i zebrafiskens rygmarv, hvor nogle axoner bruger synaptisk vesikelfrigivelse til at regulere myelinskedernes antal og længde, mens andre ikke gør det (Koudelka et al., 2016). Dette rejser den fascinerende hypotese, at kun nogle axoner er i stand til at regulere myelin via aktivitetsrelaterede signaler. Hughes et al. (2018) fandt, at deres sensoriske stimulationsparadigme ikke øgede andelen af dynamiske skeder i den somatosensoriske cortex. En mere detaljeret analyse af axon-subtypediversitet kombineret med en longitudinal undersøgelse af kappelængdedynamikken kunne imidlertid bekræfte, om kappelængde-remodellering er specifik for visse kredsløb.
Spejler kappelængdestabilitet en manglende evne til at remodellere kapper? Eksperimenter i zebrafisk tyder på, at remodellering af kappelængden kan induceres, når myeliniseringsprofilen af et axon forstyrres. Auer et al. (2018) ablated enkelte oligodendrocytter og derfor sparsomt fjernede kapper langs axoner. De fandt, at når en enkelt myelinskede tabes på et fuldt myeliniseret axon, kunne de tilstødende skeder genstarte hurtig vækst for at dække det umyeliniserede hul. I flere tilfælde blev der dannet en ny myelinskede i stedet for dens forgænger og kunne endda skubbe sig tilbage mod de invaderende naboskeder for at genoprette det oprindelige myeliniseringsmønster (figur 2A). Nogle gange opretholdes et specifikt myeliniseringsmønster derfor fortrinsvis, selv efter myelinafbrydelse. Dette kan være for at opretholde de optimerede ledningsegenskaber for det underliggende axon. Auer et al. (2018) observerede sparsomt myeliniserede axoner i larval zebrafisk, som tidligere identificeret i gnavercortex. Interessant nok fandt de, at ved ablation af enkelte kapper på sådanne sparsomt myeliniserede axoner blev der dannet en ny kappe på stort set samme sted som den ablatede kappe, selv langs en ellers umyeliniseret strækning af axonet (Figur 2B). Således synes myeliniseringsmønstrene langs sparsomt myeliniserede axoner også at blive stabilt opretholdt i zebrafisk, som foreslået af Hill et al. (2018) i gnavere. Funktionen af sparsomme myeliniseringsprofiler er fortsat ukendt. Sådanne mønstre kan muliggøre en mere dynamisk finjustering af enkelt axonfunktion over tid, selv om det også er muligt, at sådanne umyeliniserede huller kan lette gradvis myelinisering for at opretholde konsistente ledningstider inden for kredsløb, efterhånden som dyret vokser og/eller axonlængder ændres.
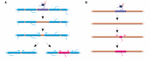
FIGUR 2. Myelinomdannelse kan forekomme in vivo. (A) Ablation af enkelte kapper på et fuldt myeliniseret axon kan fremkalde hurtig vækst af naboskakter til at dække hullet. Dette hul kan enten dækkes helt af de tilstødende kapper, eller den oprindelige myeliniseringsprofil kan genoprettes ved tilføjelse af en ny kappe. (B) Ablation af en kappe på et sparsomt myeliniseret axon efterfølges af dannelse af en ny myelinkappe af identisk størrelse og placering som den ablatede forgængerkappe. Sammenfatning af data fra Auer et al. (2018).
Har stabile myelinskeder hos pattedyr også denne evne til at remodelleres, når myeliniseringsmønsteret forstyrres? Der er behov for yderligere longitudinale undersøgelser kombineret med demyelinisering for at besvare dette spørgsmål. Det er muligt, at en sådan remodellering ikke induceres af neuronal aktivitet, men er en kompenserende mekanisme for myelintab. Aldersbetinget tab af oligodendrocytter kan udløse en remodellering af de overlevende kapper til at dække de afstødte dele af axonet og dermed bidrage til at opretholde kredsløbsfunktionen. Live-billeddannelse af myelinskeder i alderdommen kunne afgøre, om dette er tilfældet.
De live-billeddannelsesundersøgelser, der er diskuteret her, har alle vurderet myelinskedernes længde-dynamik, men ikke myelinskedernes tykkelse. Kan skindtykkelse reguleres dynamisk? Stimulering af PI3K/AKT/mTOR-signalering i oligodendrocytter hos voksne mus udløser yderligere myelinindpakning for at øge skindtykkelsen (Snaidero et al., 2014). Dette kan moduleres af kredsløbsaktivitet, da neuronal stimulering fører til øget kappetykkelse i både juvenile og voksne mus (Gibson et al., 2014; Mitew et al., 2018). Dette fremhæver behovet for at afbilde alle skedeparametre longitudinalt for fuldt ud at forstå dynamikken i skedeomdannelsen. Der er behov for live-billeddannelsesmodaliteter til nøjagtig måling af kappens tykkelse langs axoner, da dette i øjeblikket kræver tværsnitsmåling via elektronmikroskopi, hvilket begrænser analysen til et enkelt tidspunkt. Nogle mærkningsfrie billeddannelsesteknikker, såsom tredje harmonisk generationsmikroskopi og spektralreflektometri, viser løfter til at udføre sådanne målinger (Lim et al., 2014; Kwon et al., 2017). Kobling af disse teknikker med longitudinelle undersøgelser af gnavercortex kunne bestemme, om etablerede myelinskeder kan justere deres tykkelse, eller om neuronal aktivitet simpelthen skubber de novo myelinisering til at producere tykkere skeder.
Det ser derfor ud til, at selvom myelinskeder er i stand til at remodelleres, når myelin er forstyrret, er de fleste skeder generelt stabile i længden. Denne stabilitet skyldes muligvis opretholdelse af tidligt etablerede myeliniseringsmønstre, der er optimeret til kredsløbsfunktion.
Fremtiden
Nylige billeddannelsesundersøgelser af pattedyr har fokuseret på de novo myelinisering og remodellering af kappe i kortikal grå substans. Kortikale kredsløb modtager og sender information via mange regioner, såsom rygmarven og corpus callosum, og derfor kan ændringer i myelin i flere forskellige områder af CNS ændre signaleringen i et enkelt kredsløb. CNS beskrives traditionelt efter udseende efter formaldehydfiksering, hvor “hvid substans” beskriver de stærkt myeliniserede axonbaner, mens “grå substans” beskriver regioner, der er tæt pakket med neuronale cellekroppe, dendritter og synapser. Denne klassifikation er imidlertid for forenklet; OPC’er producerer myeliniserende oligodendrocytter i både grå og hvid substans (Dawson et al., 2003), og der er faktisk nye beviser for diversitet i den oligodendrogliale slægt og i mønstre af myelinisering i både grå og hvid substans (Rivers et al., 2008; Viganò et al., 2013; Young et al., 2013; Bechler et al., 2015). En sådan diversitet kan afspejle de unikke krav til myelin i forskellige områder, og potentielt på forskellige kredsløb, i CNS. Yderligere longitudinale billeddannelsesundersøgelser er nødvendige for bedre at forstå dynamikken af de novo myelinisering og sheath remodellering i områder af CNS ud over cortex.
Mens den optiske gennemsigtighed i larvezebrafiskens optiske gennemsigtighed egner sig til ikke-invasiv levende billeddannelse, er det mere invasivt og teknisk udfordrende at udføre sådanne eksperimenter i pattedyrs CNS. Hill et al. (2018) og Hughes et al. (2018) udnyttede to-fotonmikroskopi med kraniale billedvinduer til at afbilde dybder op til 400 μm ind i cortexen. Lignende teknikker kunne bruges til at afbilde overfladiske myeliniserede baner i rygmarven over tid (Locatelli et al., 2018), men dybere CNS-regioner kan ikke gennemtrænges af to-fotonmikroskopi alene. Et alternativ er at bruge to-foton mikroendoskopi, hvor en mikroendoskopisk sonde med en gradient refraktivt indeks (GRIN) linse indsættes i vævet for at afbilde celler dybere i hjernen . Indføring af endoskopet kan dog føre til inflammatoriske reaktioner, som kan påvirke myeliniseringen. Et alternativ kunne være tre-fotonmikroskopi ved hjælp af den kraniale billedvinduesmetode, som også tidligere er blevet brugt til at afbilde hippocampus (Horton et al., 2013; Ouzounov et al., 2017). Tre-fotonmikroskopi giver et betydeligt større signal-til-baggrund-forhold end to-fotonmikroskopi og kan derfor bruges til at afbilde dybere vævsstrukturer.
Det er særligt vigtigt at overveje ikke kun forskellige CNS-regioner, men også forskellige neuroner inden for disse regioner. Tidligere forskning tyder på, at der er mekanistiske forskelle i, hvordan forskellige neuronsubtyper regulerer deres myelinisering (Koudelka et al., 2016). Derudover kan der være diversitet i den lokale regulering af myelin. Det er vigtigt at huske, at forskellige dele af CNS ikke er separate enheder, men er indbyrdes forbundne. Integration af mesoskala connectomics, som fokuserer på at forstå forbindelserne mellem forskellige neuronsubtyper på tværs af forskellige regioner (Zeng, 2018), vil være afgørende for vores forståelse af, hvordan den livslange myeliniseringsdynamik varierer mellem forskellige kredsløb.
Hvad er den funktionelle konsekvens af myelinregulering langs forskellige kredsløb? Indtil videre kan de funktionelle konsekvenser kun udledes af korrelationer med adfærd. I sidste ende er der behov for at koble måling af myelin-dynamikken med direkte vurdering af kredsløbsaktivitet. Dette vil kræve registrering af neuronal aktivitet under longitudinelle undersøgelser af myelinisering for direkte at forbinde de novo myelinisering eller remodellering af skeden med ændringer i kredsløbsfunktionen over tid. Det vil være vigtigt at måle myelin-dynamikken og den elektrofysiologiske aktivitet af individuelle neuroner og axoner for at fastslå, hvordan ændringer i de forskellige myelinskedeparametre rent faktisk påvirker ledningsegenskaberne på enkeltcelleniveau, samt at vurdere aktiviteten på populationsniveau. Værktøjer som genetisk kodede Ca2+- eller spændingsindikatorer muliggør relativt ikke-invasiv registrering af kredsløbsaktivitet og kan endda bruges til at vurdere kredsløbsaktivitet i hele hjernen (Ahrens et al., 2012; Lovett-Barron et al., 2017).
Konklusion
Myelinisering af axoner repræsenterer en kraftfuld potentiel mekanisme til regulering af kredsløbsfunktion gennem hele livet. Forskning har vist, at de novo myelinisering i cortex (via produktion af nye oligodendrocytter) forekommer selv i voksenalderen, og at dette kan forstærkes ved at stimulere kredsløbsaktivitet. Når først myelin er dannet, er det stabilt med en lille udskiftning af oligodendrocytter og begrænset remodellering af længden af de eksisterende myelinskeder. Disse stabile strukturer kan dog bevare evnen til at omforme sig, hvis myelinet forstyrres. Dette har interessante implikationer for myelins plasticitet med hensyn til at opretholde kredsløbsfunktionen under skader, sygdom og alderdom. Det er endnu uvist, hvordan ændringer i myeliniseringen påvirker funktionen af det underliggende kredsløb. I sidste ende er der behov for en tilgang på kredsløbsniveau, der integrerer analyse af myelinets dynamik med direkte måling af kredsløbsfunktionen, for fuldt ud at kunne vurdere, hvordan dynamisk myelinisering påvirker nervesystemets samlede funktion gennem hele livet.
Author Contributions
Alle forfattere på listen har ydet et væsentligt, direkte og intellektuelt bidrag til arbejdet og godkendt det til offentliggørelse.
Funding
DAL er støttet af et Wellcome Trust Senior Research Fellowship (102836/Z/13/Z). JMW er støttet af et Ph.D. Tissue Repair Studentship Award fra University of Edinburgh (MRC Doctoral Training Partnership MR/K501293/1) og Wellcome Trust Four-Year Ph.D. Program in Tissue Repair (Grant 108906/Z/15/Z).
Interessekonflikterklæring
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller finansielle relationer, der kunne opfattes som en potentiel interessekonflikt.
Ahrens, M. B., Li, J. M. M., Orger, M. B., Robson, D. N., Schier, A. F., Engert, F., et al. (2012). Hjernedækkende neuronal dynamik under motorisk tilpasning i zebrafisk. Nature 485, 471-477. doi: 10.1038/nature11057
PubMed Abstract | CrossRef Full Text | Google Scholar
Almeida, R. G., and Lyons, D. A. (2017). Om myeliniseret axonplasticitet og neuronal kredsløbsdannelse og funktion. J. Neurosci. 37, 10023-10034. doi: 10.1523/JNEUROSCI.3185-16.2017
CrossRef Full Text | Google Scholar
Arancibia-Cárcamo, I. L., Ford, M. C., Cossell, L., Ishida, K., Tohyama, K., og Attwell, D. (2017). Node of Ranvier length as a potential regulator of myelinated axon conduction speed. eLife 6:e23329. doi: 10.7554/eLife.23329
PubMed Abstract | CrossRef Full Text | Google Scholar
Auer, F., Vagionitis, S., and Czopka, T. (2018). Bevis for myelinskedeomdannelse i CNS afsløret ved in vivo billeddannelse. Curr. Biol. 28, 549-559. doi: 10.1016/j.cub.2018.01.017
PubMed Abstract | CrossRef Full Text | Google Scholar
Barres, B. A., Hart, I. K., Coles, H. S. R., Burne, J. F., Voyvodic, J. T., Richardson, W. D., et al. (1992). Celledød og kontrol af celleoverlevelse i oligodendrocytlinjen. Cell 70, 31-46. doi: 10.1016/0092-8674(92)90531-G
CrossRef Full Text | Google Scholar
Bechler, M. E., Byrne, L., and Ffrench-Constant, C. (2015). CNS myelinskedenes længde er en iboende egenskab af oligodendrocytter. Curr. Biol. 25, 2411-2416. doi: 10.1016/j.cub.2015.07.056
PubMed Abstract | CrossRef Full Text | Google Scholar
Chang, A., Nishiyama, A., Peterson, J., Prineas, J., og Trapp, B. D. (2000). NG2-positive oligodendrocyt-progenitorceller i voksen menneskelig hjerne og multiple skleroselæsioner. J. Neurosci. 20, 6404-6412. doi: 10.1523/JNEUROSCI.20-17-06404.2000
PubMed Abstract | CrossRef Full Text | Google Scholar
Cox, S. R., Ritchie, S. J., Tucker-Drob, E. M., Liewald, D. C., Hagenaars, S. P., Davies, G., et al. (2016). Aldring og hjernens hvide stofstruktur i 3,513 UK Biobank-deltagere. Nat. Commun. 7:13629. doi: 10.1038/ncomms13629
PubMed Abstract | CrossRef Full Text | Google Scholar
Czopka, T., Ffrench-Constant, C., og Lyons, D. A. (2013). Individuelle oligodendrocytter har kun et par timer til at generere nye myelinskeder in vivo. Dev. Cell 25, 599-609. doi: 10.1016/j.devcel.2013.05.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Dawson, M. R. L., Polito, A., Levine, J. M., og Reynolds, R. (2003). NG2-udtrykkende gliale forstadieceller: en rigelig og udbredt population af cyklende celler i det voksne rotte CNS. Mol. Cell. Neurosci. 24, 476-488. doi: 10.1016/S1044-7431(03)00210-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Fard, M. K., Van der Meer, F., Sánchez, P., Cantuti-Castelvetri, L., Mandad, S., Jäkel, S., et al. (2017). BCAS1-ekspression definerer en population af tidlige myeliniserende oligodendrocytter i multipel sklerose-læsioner. Sci. Transl. Med. 9:eaam7816. doi: 10.1126/scitranslmed.aam7816
PubMed Abstract | CrossRef Full Text | Google Scholar
Fields, R. D. (2015). En ny mekanisme for nervesystemets plasticitet: aktivitetsafhængig myelinisering. Nat. Rev. Neurosci. 16, 756-767. doi: 10.1038/nrn4023
PubMed Abstract | CrossRef Full Text | Google Scholar
Ford, M. C., Alexandrova, O., Cossell, L., Stange-Marten, A., Sinclair, J., Kopp-Scheinpflug, C., et al. (2015). Tuning af Ranvier node og internode egenskaber i myelinerede axoner for at justere aktionspotentiale timing. Nat. Commun. 6:8073. doi: 10.1038/ncomms9073
PubMed Abstract | CrossRef Full Text | Google Scholar
Gibson, E. M., Purger, D., Mount, C. W., Goldstein, A. K., Lin, G. L., Wood, L. S., et al. (2014). Neuronal aktivitet fremmer oligodendrogenese og adaptiv myelinisering i pattedyrhjernen. Science 344:1252304. doi: 10.1126/science.1252304
PubMed Abstract | CrossRef Full Text | Google Scholar
Hill, R. A., Li, A. M., and Grutzendler, J. (2018). Livslang kortikal myelinplasticitet og aldersrelateret degeneration i den levende pattedyrhjerne. Nat. Neurosci. 21, 683-695. doi: 10.1038/s41593-018-0120-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Hill, R. A., Patel, K. D., Goncalves, C. M., Grutzendler, J., og Nishiyama, A. (2014). Modulation af oligodendrocytgenerering i et kritisk tidsmæssigt vindue efter NG2-celledeling. Nat. Neurosci. 17, 1518-1527. doi: 10.1038/nn.3815
PubMed Abstract | CrossRef Full Text | Google Scholar
Horton, N. G., Wang, K., Kobat, D., Clark, C. G., Wise, F. W., Schaffer, C. B., et al. (2013). In vivo tre-fotonmikroskopi af subkortikale strukturer i en intakt musehjerne. Nat. Photonics 7, 205-209. doi: 10.1038/NPHOTON.2012.336
PubMed Abstract | CrossRef Full Text | Google Scholar
Huang, B., Wei, W., Wang, G., Gaertig, M. A., Feng, Y., Wang, W., et al. (2015). Mutant huntingtin nedregulerer myelinreguleringsfaktor-medieret myelin-genekspression og påvirker modne oligodendrocytter. Neuron 85, 1212-1226. doi: 10.1016/j.neuron.2015.02.026
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, E. G., Kang, S. H., Fukaya, M., og Bergles, D. E. (2013). Oligodendrocyte progenitors balancerer vækst med selvafvisning for at opnå homeostase i den voksne hjerne. Nat. Neurosci. 16, 668-676. doi: 10.1038/nn.3390
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., og Bergles, D. E. (2018). Myelinomformning gennem erfaringsafhængig oligodendrogenese i den voksne somatosensoriske cortex. Nat. Neurosci. 21, 696-706. doi: 10.1038/s41593-018-0121-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Jung, J. C., Mehta, A. D., Aksay, E., Stepnoski, R., og Schnitzer, M. J. (2004). In vivo billeddannelse af pattedyrs hjerne ved hjælp af en- og to-foton fluorescensmikroendoskopi. J. Neurophysiol. 92, 3121-3133. doi: 10.1152/jn.00234.2004
PubMed Abstract | CrossRef Full Text | Google Scholar
Kang, S. H., Li, Y., Fukaya, M., Lorenzini, I., Cleveland, D. W., Ostrow, L. W., et al. (2013). Degeneration og nedsat regeneration af grå substans oligodendrocytter i amyotrofisk lateral sklerose. Nat. Neurosci. 16, 571-579. doi: 10.1038/nn.3357
PubMed Abstract | CrossRef Full Text | Google Scholar
Koudelka, S., Voas, M. G., Almeida, R. G., Baraban, M., Soetaert, J., Meyer, M. P., et al. (2016). Individuelle neuronale undertyper udviser diversitet i CNS-myelinisering medieret af synaptisk vesikelfrigivelse. Curr. Biol. 26, 1447-1455. doi: 10.1016/j.cub.2016.03.070
PubMed Abstract | CrossRef Full Text | Google Scholar
Kougioumtzidou, E., Shimizu, T., Hamilton, N. B., Tohyama, K., Sprengel, R., Monyer, H., et al. (2017). Signalering gennem AMPA-receptorer på oligodendrocytprækursorer fremmer myelinisering ved at øge oligodendrocytoverlevelsen. eLife 6:e2808080. doi: 10.7554/eLife.28080
PubMed Abstract | CrossRef Full Text | Google Scholar
Kwon, J., Kim, M., Park, H., Kang, B.-M., Jo, Y., Kim, J.-H., et al. (2017). Etiketfri nanoskala optisk metrologi på myeliniserede axoner in vivo. Nat. Commun. 8:1832. doi: 10.1038/s41467-017-01979-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Levene, M. J., Dombeck, D. A., Kasischke, K. A., Molloy, R. P., og Webb, W. W. (2004). In vivo multiphotonmikroskopi af dybt hjernevæv. J. Neurophysiol. 91, 1908-1912. doi: 10.1152/jn.01007.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Lim, H., Sharoukhov, D., Kassim, I., Zhang, Y., Salzer, J. L., og Melendez-Vasquez, C. V. (2014). Mærkningsfri billeddannelse af Schwann-celle myelination ved tredje harmonisk generationsmikroskopi. Proc. Natl. Acad. Sci. U.S.A. 111, 18025-18030. doi: 10.1073/pnas.1417820111
PubMed Abstract | CrossRef Full Text | Google Scholar
Locatelli, G., Theodorou, D., Kendirli, A., Jordão, M. J. C., Jordão, M. J. C., Staszewski, O., Phulphagar, K., et al. (2018). Mononukleære fagocytter specificerer og tilpasser lokalt deres fænotype i en multipel sklerosemodel. Nat. Neurosci. 21, 1196-1208. doi: 10.1038/s41593-018-0212-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovett-Barron, M., Andalman, A. S., Allen, W. E., Vesuna, S., Kauvar, I., Burns, V. M., et al. (2017). Ancistrale kredsløb til koordineret modulering af hjernens tilstand. Cell 171, 1411-1423. doi: 10.1016/j.cell.2017.10.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Mabbott, D. J., Noseworthy, M., Bouffet, E., Laughlin, S., og Rockel, C. (2006). Vækst i det hvide stof som en mekanisme for kognitiv udvikling hos børn. Neuroimage 33, 936-946. doi: 10.1016/j.neuroimage.2006.07.024
PubMed Abstract | CrossRef Full Text | Google Scholar
McKenzie, I. A., Ohayon, D., Li, H., Paes de Faria, J., Emery, B., Tohyama, K., et al. (2014). Motorisk færdighedsindlæring kræver aktiv central myelination. Science 346, 318-322. doi: 10.1126/science.1254960
PubMed Abstract | CrossRef Full Text | Google Scholar
Mitew, S., Gobius, I., Fenlon, L. R., McDougall, S. J., Hawkes, D., Xing, Y. L., et al. (2018). Farmakogenetisk stimulering af neuronal aktivitet øger myelinisering på en axonspecifik måde. Nat. Commun. 9:306. doi: 10.1038/s41467-017-02719-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Mount, C. W., og Monje, M. (2017). Indpakket for at tilpasse sig: erfaringsafhængig myelination. Neuron 95, 743-756. doi: 10.1016/j.neuron.2017.07.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Ouzounov, D. G., Wang, T., Wang, M., Feng, D. D., Horton, N. G., Cruz-Hernández, J. C., et al. (2017). In vivo tre-foton billeddannelse af aktivitet af GCaMP6-mærkede neuroner dybt i intakt musehjerne. Nat. Methods 14, 388-390. doi: 10.1038/nmeth.4183
PubMed Abstract | CrossRef Full Text | Google Scholar
Ritchie, S. J., Bastin, M. E., Tucker-Drob, E. M., Maniega, S. M., Engelhardt, L. E., Cox, S. R., et al. (2015). Koblede ændringer i hjernens hvide stofmikrostruktur og flydende intelligens senere i livet. J. Neurosci. 35, 8672-8682. doi: 10.1523/JNEUROSCI.0862-15.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Rivers, L. E., Young, K. M., Rizzi, M., Jamen, F., Psachoulia, K., Wade, A., et al. (2008). PDGFRA/NG2-glia genererer myeliniserende oligodendrocytter og piriforme projektionsneuroner hos voksne mus. Nat. Neurosci. 11, 1392-1401. doi: 10.1038/nn.2220
PubMed Abstract | CrossRef Full Text | Google Scholar
Sampaio-Baptista, C., Khrapitchev, A. A. A., Foxley, S., Schlagheck, T., Scholz, J., Jbabdi, S., et al. (2013). Motorisk færdighedsindlæring inducerer ændringer i hvidt stof mikrostruktur og myelinisering. J. Neurosci. 33, 19499-19503. doi: 10.1523/jneurosci.3048-13.2013
PubMed Abstract | CrossRef Full Text | Google Scholar
Scantlebury, N., Cunningham, T., Dockstader, C., Laughlin, S., Gaetz, W., Rockel, C., et al. (2014). Forhold mellem modning af hvidt stof og reaktionstid i barndommen. J. Int. Neurophsychol. Soc. 20, 99-112. doi: 10.1017/S1355617713001148
PubMed Abstract | CrossRef Full Text | Google Scholar
Schain, A. J., Hill, R. A., og Grutzendler, J. (2014). Mærkningsfri in vivo billeddannelse af myeliniserede axoner i sundhed og sygdom med spektral konfokal refleksionsmikroskopi. Nat. Med. 20, 443-449. doi: 10.1038/nm.3495
PubMed Abstract | CrossRef Full Text | Google Scholar
Scholz, J., Klein, M. C., Behrens, T. E. J., og Johansen-berg, H. (2009). Træning inducerer ændringer i den hvide stofs arkitektur. Nat. Neurosci. 12, 1370-1371. doi: 10.1038/nn.2412
PubMed Abstract | CrossRef Full Text | Google Scholar
Snaidero, N., Möbius, W., Czopka, T., Hekking, L. H. P., Mathisen, C., Verkleij, D., et al. (2014). Myelinmembranindpakning af CNS axoner ved PI(3,4,5,5)P3-afhængig polariseret vækst ved den indre tunge. Cell 156, 277-290. doi: 10.1016/j.cell.2013.11.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Takahashi, N., Sakurai, T., Davis, K. L., og Buxbaum, J. D. (2011). Sammenkobling af oligodendrocyt- og myelin-dysfunktion til neurokredsløbsmæssige abnormiteter i skizofreni. Prog. Neurobiol. 93, 13-24. doi: 10.1016/j.pneurobio.2010.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Tomassy, G. S., Berger, D. R., Chen, H.-H., Kasthuri, N., Hayworth, K. J., Vercelli, A., et al. (2014). Distinkte profiler af myelinfordeling langs enkelte axoner af pyramidale neuroner i neocortex. Science 344, 319-324. doi: 10.1126/science.1249766
PubMed Abstract | CrossRef Full Text | Google Scholar
Tripathi, R. B., Jackiewicz, M., Mckenzie, I. A., I. A., Kougioumtzidou, E., Grist, M., and Richardson, W. D. (2017). Bemærkelsesværdig stabilitet af myeliniserende oligodendrocytter i mus. Cell Rep. 21, 316-323. doi: 10.1016/j.celrep.2017.09.050
PubMed Abstract | CrossRef Full Text | Google Scholar
Viganò, F., Möbius, W., Götz, M., og Dimou, L. (2013). Transplantation afslører regionale forskelle i oligodendrocytdifferentiering i den voksne hjerne. Nat. Neurosci. 16, 1370-1372. doi: 10.1038/nn.3503
PubMed Abstract | CrossRef Full Text | Google Scholar
Watkins, T. A., Emery, B., Mulinyawe, S., og Barres, B. A. (2008). Distinkte stadier af myelinisering reguleret af gamma-sekretase og astrocytter i et hurtigt myeliniserende CNS-kokultursystem. Neuron 60, 555-569. doi: 10.1016/j.neuron.2008.09.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Waxman, S. G. (1980). Bestemmelser af ledningshastighed i myeliniserede nervefibre. Muscle Nerve 3, 141-150. doi: 10.1002/mus.88003030207
PubMed Abstract | CrossRef Full Text | Google Scholar
Xiao, L., Ohayon, D., Mckenzie, I. A., Sinclair-Wilson, A., Wright, J. L., Fudge, A. D., et al. (2016). Hurtig produktion af nye oligodendrocytter er nødvendig i de tidligste faser af indlæring af motoriske færdigheder. Nat. Neurosci. 19, 1210-1217. doi: 10.1038/nn.4351
PubMed Abstract | CrossRef Full Text | Google Scholar
Yeung, M. S. Y., Zdunek, S., Bergmann, O., Bernard, S., Salehpour, M., Alkass, K., et al. (2014). Dynamikken af oligodendrocytgenerering og myelinisering i den menneskelige hjerne. Cell 159, 766-774. doi: 10.1016/j.cell.2014.10.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Young, K. M., Psachoulia, K., Tripathi, R. B., Dunn, S.-J., Cossell, L., Attwell, D., et al. (2013). Oligodendrocytdynamik i det sunde voksne CNS: beviser for myelinomdannelse. Neuron 77, 873-885. doi: 10.1016/j.neuron.2013.01.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Zeng, H. (2018). Mesoskala connectomics. Curr. Opin. Neurobiol. 50, 154-162. doi: 10.1016/j.conb.2018.03.003
PubMed Abstract | CrossRef Full Text | Google Scholar