
Avant l’attaque des chenilles : Semis de chou rouge russe
Jeanne transforme sa frustration face aux chenilles dans son jardin en une exploration de la botanique derrière une espèce extraordinaire : Brassica oleracea.
Les papillons blancs du chou (Lepidoptera : Pieris rapae) ont décimé la culture automnale de chou frisé dans notre jardin. Pour être juste, les chenilles vertes abondantes n’ont pas consommé le limbe entier de chaque feuille. Les feuilles grignotées restantes, cependant, selon mon mari, ne ressemblaient plus tant à de la nourriture qu’à un élevage de chenilles qu’il serait fastidieux de transformer en nourriture. Il a arraché l’élevage de chenilles, l’a jeté dans le bac à compost et l’a remplacé par de la laitue. Contrairement au chou frisé, qui fait partie de la famille de la moutarde (Brassicaceae), la laitue fait partie de la famille des tournesols (Asteraceae) et n’est donc pas du tout attrayante pour les papillons du chou blanc.

Dégâts de chenilles sur de jeunes choux verts frisés dans le jardin de Monticello
J’ai été tenté de sauver les feuilles criblées de trous de leur destin de compost, en partie parce que je sais que la mastication des chenilles a en fait augmenté la concentration foliaire de certains des composés qui contribuent à la réputation nutritive du chou frisé, et aussi parce que la chute des températures en automne confère une douceur extraordinaire au chou frisé et aux autres légumes crucifères qui sont en fait tous des variétés de la même espèce, Brassica oleracea : le chou-fleur, le chou, le chou-rave, le chou de Bruxelles, le brocoli chinois et le chou-fleur. Les détails des conséquences chimiques de la consommation de chenilles feront bientôt l’objet d’un article à part entière. Ce post est consacré à la botanique et à la biologie de l’évolution derrière l’étonnante diversité de B. oleracea.
Sélection artificielle et Brassica oleracea

Fleurs de moutarde jaune sauvage (Sinapis arvensis)
Dans son ouvrage De l’origine des espèces au moyen de la sélection naturelle, Charles Darwin établit un parallèle entre la sélection naturelle dans la nature et l’élevage de plantes et d’animaux orchestré par les agriculteurs, une pratique englobée par ce que les biologistes appellent la « sélection artificielle » : « Si l’homme peut par patience sélectionner les variations qui lui sont utiles, pourquoi […] les variations utiles aux produits vivants de la nature ne se présenteraient-elles pas souvent, et ne seraient-elles pas préservées et sélectionnées ? » La sélection naturelle est ce que l’on appelle communément « la survie des plus aptes » : la survie et le succès reproductif disproportionnés de certains individus dans une population, souvent ceux dont les caractéristiques particulières les rendent mieux adaptés à leur environnement que leurs congénères moins chanceux. Dans ce contexte, l' »environnement » englobe non seulement les conditions abiotiques subies par un organisme, telles que le climat et le sol dominants, mais aussi les autres organismes avec lesquels il interagit, y compris ses concurrents, ses prédateurs, les agents pathogènes et, dans le cas de certaines plantes, ses pollinisateurs et ses disperseurs de fruits.

Chou de Savoie rouge, commençant tout juste à développer une tête
La sélection artificielle exige qu’un agriculteur, en plus des circonstances environnementales naturelles, détermine quels individus possèdent les caractéristiques clés à transmettre à la génération suivante, en permettant seulement aux individus les plus désirables de se reproduire. L’espoir est que le trait particulier faisant l’objet de la sélection soit hautement héritable (ayant une forte base génétique, plutôt qu’environnementale), de sorte que la génération suivante aura une plus grande expression du trait que la génération parentale.

Broccoli
Les agriculteurs ont itéré ce processus pendant plus de 10 000 ans, depuis que les premiers agriculteurs ont pris les graines des arbres fruitiers les plus juteux et des tiges de céréales les plus lourdement chargées et les ont plantées près de leurs maisons. Ce faisant, nos ancêtres agriculteurs ont domestiqué un grand nombre des espèces de plantes sauvages les plus importantes sur le plan nutritionnel et les plus délicieuses, les transformant au fil des générations en variétés que les hommes modernes reconnaissent comme les principales plantes alimentaires. Les agriculteurs choisissent d’élever les plantes individuelles qui maximisent les qualités souhaitables et minimisent les indésirables. Dans la variété domestiquée d’une espèce sauvage, la qualité souhaitable faisant l’objet de la sélection, comme la douceur du fruit ou la taille d’une structure particulière, peut être amplifiée avec le temps, exagérée par rapport à la condition sauvage moyenne. Ou un agriculteur peut sauver de l’obscurité un individu avec une caractéristique rare dans la nature ou dans la variété cultivée la plus commune, comme une couleur de fleur particulière, ou une amande mutante non toxique, faisant de la plante unique un géniteur d’une nouvelle variété.

Têtes de brocoli et choux rouges et verts sur un marché fermier
Certaines espèces ont subi le processus de domestication plusieurs fois, et avec certaines de ces espèces, chaque effort de domestication s’est concentré sur l’amplification de différentes structures de la plante, produisant une corne d’abondance de légumes ou de fruits extraordinairement différents à partir du même progéniteur sauvage. C’est le cas de Brassica oleracea. La plante sauvage est une petite herbe qui préfère pousser sur les affleurements calcaires tout autour de la région côtière méditerranéenne. C’est une plante bisannuelle qui utilise les réserves alimentaires stockées pendant l’hiver dans sa rosette de feuilles pour produire un épi de quelques fleurs jaunes à la fin de son deuxième été avant de mourir. Ces feuilles nutritives font de ses dérivés domestiqués des cultures alimentaires importantes dans une grande partie du monde aujourd’hui. Au cours des derniers millénaires, des agriculteurs entreprenants ont contribué à la domestication de plusieurs lignées distinctes de B. oleracea, chacune amplifiant différentes parties de cette plante sauvage pour produire plusieurs variétés de légumes, ou groupes de cultivars ou sous-espèces ( » ssp. « ) : le chou frisé et le chou vert. ») : chou frisé et chou-navet (ssp. acephala), brocoli chinois (ssp. alboglabra), choux rouges et verts (ssp. capitata), chou de Milan (ssp. sabauda), chou-rave (ssp. gongylodes), chou de Bruxelles (ssp. gemmifera), brocoli (ssp. italica) et chou-fleur (ssp. botrytis). Ces variétés ont l’air dramatiquement – parfois comiquement – différentes mais sont néanmoins considérées comme la même espèce car elles sont toujours interfertiles, capables de s’accoupler entre elles et de produire une descendance fertile.

Les cultivars de Brassica oleracea sont les vedettes d’un exercice de sélection artificielle en laboratoire. Dans le sens des aiguilles d’une montre, à partir de la gauche : brocoli, chou-fleur, chou frisé décoratif, chou frisé Lacinato, chou rouge, chou de Bruxelles, chou-rave
On peut facilement observer que ces légumes ont simplement l’air très différents à première vue, de sorte qu’il est souvent surprenant pour les gens d’apprendre qu’ils sont en fait la même espèce. Pour comprendre les détails de la domestication des cultivars de B. oleracea et les structures que vous regardez lorsque vous les voyez à l’épicerie, vous devez comprendre la morphologie et le modèle de croissance de base des angiospermes (plantes à fleurs).
Brièvement, ce sont les parties qui ont été élargies dans divers cultivars :
Les feuilles – Le chou frisé, les feuilles de chou vert, Brocoli chinois
Bourgeon terminal – chou
Bourgeons axillaires (latéraux) – choux de Bruxelles
Tige – chou-rave
Inflorescences – brocoli et chou-fleur
Le corps de la plante
Les structures aériennes importantes du corps de la plante sont indiquées dans le schéma du modèle de croissance de la plante à fleurs ci-dessous.
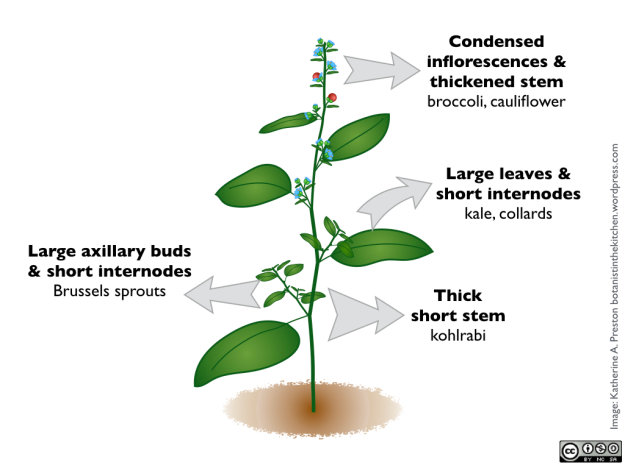
La relation entre un corps végétal générique et certaines cultures végétales. Par exemple, les variétés de Brassica oleracea résultent d’une sélection visant à exagérer différentes parties de la plante sauvage d’origine. Notez que cette plante générique n’est pas censée représenter une espèce particulière, et qu’elle ne ressemble pas à Brassica.
Le tissu du méristème chez les plantes est analogue au tissu des cellules souches chez les animaux. Il est capable de se développer en de nombreux types de structures végétales. Tous les tissus végétaux se développent par division cellulaire et ont pour origine le tissu du méristème. Lorsqu’une cellule de méristème se divise pour produire une structure particulière, comme une feuille ou une tige, seule une des deux cellules filles se différencie pour devenir une partie du corps de la plante. Cette cellule peut se diviser par la suite, mais elle ne produira que des cellules plus spécialisées, comme les feuilles ou les tiges. L’autre cellule fille reste un tissu de méristème. Les tissus de méristème situés dans différentes parties d’une plante en croissance sont nommés en fonction des structures qu’ils produisent. Le méristème des pousses, l’amas de cellules de méristème à l’extrémité de la tige principale, donne naissance aux structures végétatives (non reproductives) initiales du corps végétal en croissance.
Les plantes ont une construction modulaire et croissent en ajoutant de manière itérative des unités végétatives de base successives. Cette unité de base du corps végétal est constituée d’une tige, d’une feuille et d’un bourgeon axillaire végétatif, situé entre la tige et le pétiole d’une feuille (le pédoncule en forme de tige qui attache le limbe de la feuille à la tige). Le point de la tige où la feuille et le bourgeon axillaire qui lui est associé prennent naissance est un nœud. Au fur et à mesure de la croissance de la plante, le méristème des pousses continue à générer l’unité de base, espacée sur la tige d’une longueur d’entre-nœud déterminée. Les bourgeons axillaires contiennent eux-mêmes un tissu de méristème, appelé méristème axillaire, qui peut donner naissance à des branches latérales de la tige principale, qui elles-mêmes répéteront l’unité végétative de base.
Choux frisé, chou vert et brocoli chinois : expansion des feuilles

Choux frisé russe rouge et brocoli chinois
L’histoire précoce de B.oleracea n’est pas bien connue, mais elle était déjà bien établie comme légume de jardin à l’époque où l’ancien Grec Théophraste (371-287 av. J.-C.) a écrit son Enquête sur les plantes, dans laquelle il mentionne trois variétés de feuilles, comme le chou frisé et les feuilles de chou vert, et enregistre la croyance dominante à l’époque que la culture de B. oleracea à proximité des raisins donnerait une saveur de chou au vin. La domestication a augmenté la taille des feuilles de B. oleracea sauvage pour produire le chou frisé, les feuilles de chou cavalier en Europe et le brocoli chinois en Chine.

Choux frisé vert et feuilles de chou cavalier
Ces variétés feuillues sont les plus similaires à (les « moins dérivées » de) l’ancêtre sauvage. Le chou frisé et le chou cavalier étaient probablement les progéniteurs de toutes les autres variétés (choux de Bruxelles, choux, choux-raves, et brocoli et chou-fleur en Europe ; brocoli chinois en Asie) (Maggioni et al. 2010).
Plusieurs variétés de chou frisé sont de plus en plus disponibles sur les marchés et varient en termes de couleur, de morphologie des feuilles et de texture. Le chou vert frisé est le plus couramment disponible, suivi du chou lacinato ou dinosaure, dont les feuilles ont des fossettes vertes foncées, et des variétés russes frisées « rouges » violettes et frisées rouges et blanches. Les choux décoratifs violets, blancs et verts sont également des plantes ornementales courantes dans les jardins d’automne.
Chou : expansion du bourgeon terminal

Noyau du chou
Pour faire le saut de développement des variétés domestiques à feuilles vers le chou pommé, les agriculteurs ont conservé les grandes feuilles du chou frisé précoce et se sont concentrés sur la réduction spectaculaire de la longueur des entre-nœuds et l’élargissement de la taille du bourgeon terminal. Le bourgeon terminal est la grappe de feuilles immatures issues du tissu du méristème de la pousse. La tête du chou est un bourgeon terminal extraordinairement grand, entouré de grandes feuilles serrées les unes contre les autres sur la tige courte et grasse (le cœur de la tête du chou, illustré à droite). Trois variétés distinctes de choux sont reconnues : le blanc (B. oleracea ssp. capitata var. alba), le rouge (B. o. c. var. rubra) et le savoy (B. oleracea ssp. sabauda), qui a une tête vert pâle avec des feuilles à fossettes, semblable à la texture du chou frisé lacinato. Le chou pommé était un légume domestique reconnu en Méditerranée au moins au XIIe siècle.
Choux de Bruxelles : expansion des bourgeons axillaires

Choux de Bruxelles sur la tige, feuilles enlevées, pétiole étiqueté
Les choux de Bruxelles ont été développés par sélection pour élargir considérablement les bourgeons axillaires. Vous pouvez le constater si jamais vous les cultivez vous-même ou si vous pouvez les acheter sur la tige lorsqu’ils sont en saison à la fin de l’été et à l’automne. En général, lorsque vous achetez des choux de Bruxelles sur pied, les feuilles ont été coupées, de sorte que vous ne pouvez voir que les choux eux-mêmes, disposés en spirales serrées autour d’une tige principale épaisse. Pour savoir s’il s’agit de bourgeons axillaires, regardez les cicatrices de feuilles ou les pétioles (les « tiges » des feuilles) sous chaque chou de Bruxelles (voir la photo de droite avec le pétiole étiqueté et la cicatrice de feuille pâle visible sous le chou). Les bourgeons axillaires sont des versions plus petites du bourgeon terminal qui a été agrandi pour produire le chou, ce qui explique pourquoi les choux de Bruxelles ressemblent à des choux miniatures. Les choux de Bruxelles sont récoltés lorsque les feuilles qui se déploient sont encore bien enveloppées dans le bourgeon. Si on les laissait se développer, ils deviendraient des branches courtes et épaisses. Si vous coupez un chou de Bruxelles en son milieu, vous verrez une petite tige courte et grasse, comme un trognon de chou, et si vous regardez de très près entre cette petite tige et les feuilles qui se déploient à partir de ses petits nœuds serrés, vous verrez d’adorables petits bourgeons axillaires. Ces petits bourgeons axillaires sont d’ailleurs toujours visibles après que les choux de Bruxelles coupés en tranches ont été braisés dans un peu de bouillon et mélangés à du beurre brun dans lequel vous avez fait sauter des graines de moutarde brune (une autre espèce de Brassica et le sujet d’un prochain article) – juste au cas où vous voudriez impressionner vos amis au dîner avec vos étonnantes connaissances botaniques. Le chou de Bruxelles est populaire dans son homonyme belge et pourrait y avoir été développé vers le 13e siècle.
Le chou-rave : expansion de la tige

Le chou-rave
Le tissu du méristème est également situé sur les côtés des tiges, en plus des bourgeons terminaux et axillaires. Ce tissu de méristème latéral est ce qui permet à la tige de croître vers l’extérieur, et pas seulement vers le haut, pour soutenir la plante en croissance. Pour comprendre les origines morphologiques du chou-rave, vous pouvez commencer par imaginer le chou frisé et réfléchir aux changements nécessaires pour le transmogrifier en chou-rave. Lorsque vous achetez du chou frisé en magasin, il se présente généralement sous la forme d’un paquet de feuilles, toutes coupées de la tige principale à partir de laquelle elles ont poussé, de sorte que vous n’avez peut-être jamais envisagé la tige principale de la plante de chou frisé. Cette tige principale n’est pas aussi épaisse que le cœur du chou, mais elle peut tout de même être substantielle (voir la tige dans les images de choux de Bruxelles ci-dessus). Cette tige charnue et comestible est remplie de cellules de parenchyme. Les cellules du parenchyme stockent les nutriments et les produits chimiques synthétisés par les plantes et fournissent un support structurel. Les nouvelles cellules du parenchyme proviennent, vous l’avez deviné, du méristème du parenchyme de la tige. Imaginez maintenant que le tissu parenchyme de cet axe principal prolifère, permettant une expansion massive du rayon de la base de la tige principale, transformant la tige en un bulbe épais et trapu. C’est l’histoire de la domestication du chou-rave, enregistrée en Europe depuis le 15e siècle (Vaughan et Geissler 2009). Les feuilles du chou-rave sont souvent coupées à l’épicerie, laissant juste de petites pointes de pétiole, mais si elles sont laissées, vous pouvez immédiatement voir (et goûter) la similitude avec les feuilles vertes du chou-rave. La peau du chou-rave est coriace – parfois le méristème latéral a produit une fine couche de bois – et il faut généralement l’éplucher. Ensuite, vous pouvez soit couper le chou-rave en fines rondelles ou en julienne pour le déguster frais, soit le couper en cubes, le rôtir ou le cuire à la vapeur et l’assaisonner comme vous le souhaitez. Le chou-rave frais coupé en julienne est excellent combiné avec des pommes ou des poires et des oignons frais coupés de façon similaire et mélangé avec une vinaigrette à la moutarde pour former une salade unique.
Brocoli et chou-fleur : expansion de l’inflorescence

Plante de brocoli, tête centrale enlevée
Le brocoli à grosse tête le plus souvent cultivé aux États-Unis n’est qu’une des variétés vertigineuses de brocoli que l’on trouve en Italie, où les premières mentions écrites du légume remontent au XVIe siècle (Gray 1982). Le chou-fleur est probablement dérivé de l’une de ces variétés italiennes de brocoli (Gray 1982). Le brocoli et le chou-fleur se distinguent par leurs inflorescences (grappes de fleurs) exceptionnellement grandes, disposées en fleurons ramifiés au sommet de leur tige principale d’inflorescence épaisse (comestible) sortant de la rosette de grandes feuilles basales typique de l’espèce. Voir la photo à droite du plant de brocoli avec l’inflorescence principale (tête de brocoli) coupée. Vous pouvez voir les grandes feuilles basales qui ressemblent beaucoup à des feuilles de chou vert et les petits fleurons latéraux du brocoli qui se ramifient à partir de la tige principale coupée. Ces grandes feuilles basales, généralement plus dures, de toutes ces variétés de B. oleracea sont tout à fait comestibles. Elles s’adouciront avec un long braisage. Un délicieux hors-d’œuvre thaïlandais, cependant, utilise des feuilles de brocoli crues comme substrat pour un wrap savoureux !

Fleurs de brocoli
L’inflorescence est généralement récoltée avant que les boutons floraux n’arrivent à maturité et ne s’ouvrent en petites fleurs jaunes typiques de l’espèce. La photo de gauche montre un morceau d’inflorescence de brocoli qui a largement dépassé l’état de récolte dans un champ de brocoli. Remarquez la similitude des fleurs avec celles de la moutarde jaune sauvage ci-dessus. Parfois, les bottes de brocoli laissées trop longtemps au réfrigérateur ouvrent quelques fleurs.
Le passage de la B. oleracea sauvage au brocoli et au chou-fleur a nécessité des modifications du moment et de l’endroit de la transition de la croissance végétative au développement reproductif. La formation de structures reproductives (fleurs, fruits, graines) sur une tige particulière nécessite deux transformations du méristème, illustrées sur le diagramme schématique ci-dessus. Tout d’abord, le méristème des pousses devient le méristème des inflorescences, qui produit la structure de la tige supportant les fleurs. Cette tige supportant les fleurs, qui peut être très ramifiée, est l’inflorescence.

Inflorescence de chou-fleur, très ramifiée
Comment faire une graine de chou-fleur
Pour obtenir réellement des fleurs au sommet d’une inflorescence, le méristème d’inflorescence doit devenir un méristème floral, qui forme les structures florales. Le « caillé » blanc serré et bosselé qui forme une tête de chou-fleur résulte de la prolifération extensive de nombreux méristèmes d’inflorescence mutants au sommet des branches d’inflorescence initiales qui s’arrêtent au stade de méristème d’inflorescence. Ils ne s’allongent jamais en inflorescence ou ne produisent pas de méristème floral. Si on laisse le chou-fleur se développer au lieu de le cueillir lorsque le caillé est le plus dense, comme c’est préférable en cuisine, le caillé se détend et environ 10 % du méristème d’inflorescence continue à produire un méristème floral et finalement des fleurs, ce qui explique pourquoi nous avons des graines de chou-fleur. Le brocoli partage avec le chou-fleur la prolifération des méristèmes d’inflorescence suivie d’un arrêt du développement de l’inflorescence, bien que les méristèmes d’inflorescence du brocoli continuent à produire des méristèmes floraux et initient le développement floral avant que l’expansion de l’inflorescence ne cesse. Les mutations génétiques qui ont conduit au type d’inflorescence du brocoli et du chou-fleur sont assez bien comprises et seront expliquées dans un prochain article. Les têtes de chou-fleur sont blanches, d’ailleurs, en raison du blanchiment par les grandes feuilles basales qui s’enroulent autour du caillé, seules ou sous la direction d’un agriculteur diligent. Si les têtes ne sont pas blanchies, elles prendront une couleur jaune verdâtre pâle et pourraient être plus amères.
Diversité extraordinaire au sein d’une même espèce
Les hybrides, résultat de croisements entre deux espèces ou groupes de cultivars, entre ces variétés complètement interfécondes produisent des légumes aux caractéristiques intermédiaires entre leurs variétés parentales. Certains de ces hybrides sont de plus en plus populaires. Le brocoli est un hybride entre les groupes de cultivars Italica et Alboglabra. Le brocoli-fleur, un chou-fleur vert, et d’autres légumes colorés ressemblant à des choux-fleurs, sont des hybrides entre les groupes Italica et Botrytis. Le brocoli rabe, ou rapini, est en fait une variété d’une autre espèce, Brassica rapa, dont il sera question dans un autre billet.
Graines de Brassica oleracea
Notez que ces plusieurs variétés ont été largement produites par des modifications de structures non reproductives. Même dans le brocoli et le chou-fleur, les sélectionneurs de plantes ont ignoré les fruits et les graines de B. oleracea pendant la domestication, et ces structures sont indiscernables parmi les plusieurs légumes (graines dans des flacons illustrés à droite). Des feuilles de texture bosselée, teintées de violet, et de nombreuses variétés vertes ont été développées de la plupart de ces légumes B. oleracea, reflétant une large variation génétique dans cette constellation de plantes qui ont été collectivement soumises à la domestication depuis des millénaires.

Cauliflower au marché fermier
Toutes ces variétés ont conservé la résistance au froid qui aide la B. oleracea sauvage à affronter les chutes de gel, une caractéristique qui a sans doute contribué à leur popularité dans les régions plus fraîches de leur Europe natale d’abord, puis du reste du globe. Les amateurs de légumes crucifères des régions tempérées fraîches se réjouissent de la douceur accrue de ces légumes à l’approche des gelées de la fin de l’été et de l’automne. L’augmentation de la concentration en sucre des tissus fait en fait partie du système antigel de nombreuses plantes, qui se met en marche lorsque les températures baissent et leur permet de continuer à pousser ou à faire mûrir les graines pendant un certain temps face au gel et de se préparer à la dormance hivernale. Profitez de la douceur caractéristique des crucifères cet automne. J’espère que les chenilles l’ont fait.
Mise à jour (8 avril 2013) : Certains choux frisés, dont le chou russe rouge, sont en fait des « choux sibériens », de l’espèce Brassica napus, la même espèce que le rutabaga, au lieu de B. oleracea.
Gray, A. R. 1982. Taxonomie et évolution du brocoli (Brassica oleracea var. italic). Economic Botany 36(4) : 397-410.
Maggioni, L., R. von Bothmer, G. Poulsen, et F. Branca. 2010. Origine et domestication des cultures de choux (Brassica oleracea L.) : considérations linguistiques et littéraires. Economic Botany 64(2) : 109-123.
Singer, S., S. Deel, et D. Walser-Kuntz. « Reconstruire l’évolution du chou-fleur et du brocoli ». Collège Carleton. http://serc.carleton.edu/genomics/units/cauliflower.html.
Vaughan, J. G., et A. Geissler. 2009. The New Oxford Book of Food Plants. Oxford University Press, New York.