Acoplamiento excitación-contracción
El acoplamiento excitación-contracción es el enlace (transducción) entre el potencial de acción generado en el sarcolema y el inicio de una contracción muscular. El desencadenante de la liberación de calcio desde el retículo sarcoplásmico hacia el sarcoplasma es una señal neural. Cada fibra muscular esquelética está controlada por una neurona motora, que conduce las señales desde el cerebro o la médula espinal hasta el músculo. La zona del sarcolema de la fibra muscular que interactúa con la neurona se denomina placa terminal motora. El extremo del axón de la neurona se denomina terminal sináptico y no entra en contacto con la placa terminal motora. Un pequeño espacio llamado hendidura sináptica separa el terminal sináptico de la placa terminal motora. Las señales eléctricas viajan a lo largo del axón de la neurona, que se ramifica a través del músculo y se conecta a las fibras musculares individuales en una unión neuromuscular.
La capacidad de las células para comunicarse eléctricamente requiere que las células gasten energía para crear un gradiente eléctrico a través de sus membranas celulares. Este gradiente de carga es transportado por iones, que se distribuyen de forma diferencial a través de la membrana. Cada ion ejerce una influencia eléctrica y una influencia de concentración. Al igual que la leche acaba mezclándose con el café sin necesidad de removerlo, los iones también se distribuyen uniformemente, si se les permite hacerlo. En este caso, no se les permite volver a un estado de mezcla uniforme.
La ATPasa sodio-potasio utiliza la energía celular para mover los iones K+ dentro de la célula y los iones Na+ fuera. Esto por sí solo acumula una pequeña carga eléctrica, pero un gran gradiente de concentración. Hay mucho K+ en la célula y mucho Na+ fuera de ella. El potasio puede salir de la célula a través de los canales de K+ que están abiertos el 90% del tiempo, y lo hace. Sin embargo, los canales de Na+ rara vez están abiertos, por lo que el Na+ permanece fuera de la célula. Cuando el K+ abandona la célula, obedeciendo a su gradiente de concentración, deja una carga negativa. Así que, en reposo, hay un gran gradiente de concentración para que el Na+ entre en la célula, y hay una acumulación de cargas negativas que quedan en la célula. Este es el potencial de membrana en reposo. El potencial en este contexto significa una separación de carga eléctrica que es capaz de hacer trabajo. Se mide en voltios, al igual que una batería. Sin embargo, el potencial transmembrana es considerablemente menor (0,07 V); por lo tanto, el pequeño valor se expresa en milivoltios (mV) o 70 mV. Como el interior de una célula es negativo en comparación con el exterior, el signo menos significa el exceso de cargas negativas dentro de la célula, -70 mV.
Si un acontecimiento cambia la permeabilidad de la membrana a los iones de Na+, éstos entrarán en la célula. Eso cambiará el voltaje. Se trata de un evento eléctrico, llamado potencial de acción, que puede utilizarse como señal celular. La comunicación se produce entre los nervios y los músculos a través de los neurotransmisores. Los potenciales de acción de las neuronas provocan la liberación de neurotransmisores de la terminal sináptica a la hendidura sináptica, donde pueden difundirse a través de la hendidura sináptica y unirse a una molécula receptora en la placa terminal motora. La placa terminal motora posee pliegues de unión -pliegues en el sarcolema que crean una gran superficie para que el neurotransmisor se una a los receptores. Los receptores son en realidad canales de sodio que se abren para permitir el paso de Na+ al interior de la célula cuando reciben la señal del neurotransmisor.
La acetilcolina (ACh) es un neurotransmisor liberado por las motoneuronas que se une a los receptores de la placa terminal motora. La liberación del neurotransmisor se produce cuando un potencial de acción recorre el axón de la motoneurona, lo que provoca una alteración de la permeabilidad de la membrana terminal sináptica y una afluencia de calcio. Los iones Ca2+ permiten que las vesículas sinápticas se muevan y se unan a la membrana presináptica (en la neurona) y liberen el neurotransmisor de las vesículas en la hendidura sináptica. Una vez liberado por el terminal sináptico, la ACh se difunde a través de la hendidura sináptica hasta la placa terminal motora, donde se une a los receptores de ACh. Al unirse el neurotransmisor, estos canales iónicos se abren y los iones de Na+ cruzan la membrana hacia la célula muscular. Esto reduce la diferencia de voltaje entre el interior y el exterior de la célula, lo que se denomina despolarización. Como la ACh se une a la placa final del motor, esta despolarización se denomina potencial de placa final. A continuación, la despolarización se extiende a lo largo del sarcolema, creando un potencial de acción a medida que los canales de sodio adyacentes al lugar de despolarización inicial detectan el cambio de voltaje y se abren. El potencial de acción se desplaza por toda la célula, creando una onda de despolarización.
La acetilcolinesterasa (AChE) descompone la ACh en acetilo y colina. La AChE reside en la hendidura sináptica, descomponiendo la ACh para que no permanezca unida a los receptores de ACh, lo que provocaría una contracción muscular prolongada no deseada (Figura 6.9).
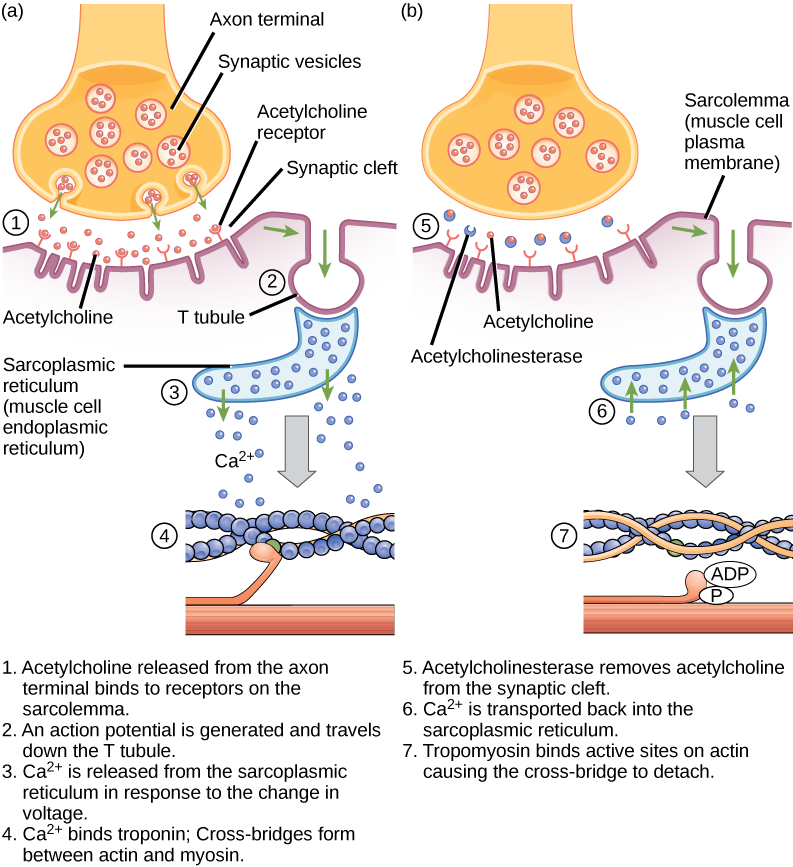
Figura 6.9. Este diagrama muestra el acoplamiento excitación-contracción en una contracción del músculo esquelético. El retículo sarcoplásmico es un retículo endoplásmico especializado que se encuentra en las células musculares.
Ejercicio \(\PageIndex{1})
El mortal gas nervioso Sarín inhibe irreversiblemente la acetilcolinesterasa. ¿Qué efecto tendría el Sarín en la contracción muscular?
Después de la despolarización, la membrana vuelve a su estado de reposo. Esto se denomina repolarización, durante la cual se cierran los canales de sodio activados por voltaje. Los canales de potasio continúan con una conductancia del 90%. Como la ATPasa de sodio-potasio de la membrana plasmática siempre transporta iones, se restablece el estado de reposo (con carga negativa en el interior en relación con el exterior). El periodo inmediatamente posterior a la transmisión de un impulso en un nervio o músculo, en el que una neurona o célula muscular recupera su capacidad para transmitir otro impulso, se denomina periodo refractario. Durante el periodo refractario, la membrana no puede generar otro potencial de acción. El periodo refractario permite que los canales iónicos sensibles al voltaje vuelvan a sus configuraciones de reposo. La ATPasa sodio-potasio mueve continuamente el Na+ hacia fuera de la célula y el K+ hacia dentro, y el K+ se escapa dejando carga negativa. Muy rápidamente, la membrana se repolariza, de modo que puede volver a despolarizarse.