Excitation-contraction coupling
Excitation-contraction coupling to związek (transdukcja) między potencjałem czynnościowym generowanym w sarkolemmie a początkiem skurczu mięśnia. Czynnikiem wyzwalającym uwalnianie wapnia z retikulum sarkoplazmatycznego do sarkoplazmy jest sygnał nerwowy. Każde włókno mięśnia szkieletowego jest kontrolowane przez neuron ruchowy, który przewodzi sygnały z mózgu lub rdzenia kręgowego do mięśnia. Obszar sarkolemmy na włóknie mięśniowym, który wchodzi w interakcję z neuronem, nazywany jest płytką końcową neuronu motorycznego. Koniec aksonu neuronu nazywany jest terminalem synaptycznym i w rzeczywistości nie styka się on z płytką motoryczną. Mała przestrzeń zwana szczeliną synaptyczną oddziela terminal synaptyczny od płytki motorycznej. Sygnały elektryczne podróżują wzdłuż aksonu neuronu, który rozgałęzia się przez mięsień i łączy się z poszczególnymi włóknami mięśniowymi w złączu nerwowo-mięśniowym.
Zdolność komórek do komunikacji elektrycznej wymaga od komórek wydatkowania energii w celu stworzenia gradientu elektrycznego w poprzek ich błon komórkowych. Ten gradient ładunku jest przenoszony przez jony, które są różnie rozmieszczone w błonie. Każdy jon wywiera wpływ elektryczny i wpływ na stężenie. Tak jak mleko w końcu zmiesza się z kawą bez konieczności mieszania, tak samo jony rozprowadzają się równomiernie, jeśli im się na to pozwoli. W tym przypadku nie wolno im powrócić do stanu równomiernego wymieszania.
ATPaza sodowo-potasowa wykorzystuje energię komórkową do przemieszczania jonów K+ wewnątrz komórki i jonów Na+ na zewnątrz. Samo to gromadzi niewielki ładunek elektryczny, ale duży gradient stężeń. W komórce znajduje się dużo K+, a na zewnątrz dużo Na+. Potas jest w stanie opuścić komórkę przez kanały K+, które są otwarte przez 90% czasu i tak się dzieje. Natomiast kanały Na+ są rzadko otwarte, więc Na+ pozostaje poza komórką. Kiedy K+ opuszcza komórkę, zgodnie z gradientem stężeń, efektywnie pozostawia za sobą ładunek ujemny. Tak więc w stanie spoczynku istnieje duży gradient stężenia dla Na+ wchodzącego do komórki, a w komórce gromadzą się ładunki ujemne. Jest to spoczynkowy potencjał membranowy. Potencjał w tym kontekście oznacza rozdzielenie ładunku elektrycznego, który jest zdolny do wykonania pracy. Jest on mierzony w woltach, tak jak w przypadku baterii. Jednak potencjał transmembranowy jest znacznie mniejszy (0,07 V); dlatego ta mała wartość jest wyrażana w miliwoltach (mV) lub 70 mV. Ponieważ wnętrze komórki jest ujemne w porównaniu z zewnętrzem, znak minus oznacza nadmiar ładunków ujemnych wewnątrz komórki, -70 mV.
Jeśli jakieś zdarzenie zmieni przepuszczalność błony dla jonów Na+, dostaną się one do komórki. To spowoduje zmianę napięcia. Jest to zdarzenie elektryczne, zwane potencjałem czynnościowym, które może być wykorzystane jako sygnał komórkowy. Komunikacja między nerwami i mięśniami odbywa się za pośrednictwem neuroprzekaźników. Potencjały czynnościowe neuronów powodują uwalnianie neuroprzekaźników z terminala synaptycznego do szczeliny synaptycznej, gdzie mogą one dyfundować przez szczelinę synaptyczną i wiązać się z cząsteczką receptora na końcowej płytce motorycznej. Płytka końcowa motoryczna posiada fałdy łączące – fałdy w sarkolemmie, które tworzą dużą powierzchnię dla neuroprzekaźnika w celu związania się z receptorami. Receptory są w rzeczywistości kanałami sodowymi, które otwierają się, aby umożliwić przejście Na+ do komórki, gdy otrzymują sygnał neuroprzekaźnika.
Acetylocholina (ACh) jest neuroprzekaźnikiem uwalnianym przez neurony ruchowe, które wiążą się z receptorami w płytce końcowej silnika. Uwolnienie neuroprzekaźnika następuje, gdy potencjał czynnościowy przemieszcza się w dół aksonu neuronu ruchowego, powodując zmianę przepuszczalności błony terminala synaptycznego i napływ wapnia. Jony Ca2+ umożliwiają pęcherzykom synaptycznym przemieszczenie się do błony presynaptycznej (na neuronie) i związanie się z nią, a następnie uwolnienie neuroprzekaźnika z pęcherzyków do szczeliny synaptycznej. Po uwolnieniu przez terminal synaptyczny, ACh dyfunduje przez szczelinę synaptyczną do końcowej płytki motorycznej, gdzie łączy się z receptorami ACh. Po związaniu neuroprzekaźnika kanały jonowe otwierają się, a jony Na+ przekraczają błonę do wnętrza komórki mięśniowej. Powoduje to zmniejszenie różnicy napięcia pomiędzy wnętrzem i zewnętrzem komórki, co nazywane jest depolaryzacją. Ponieważ ACh wiąże się z płytką końcową silnika, depolaryzacja ta nazywana jest potencjałem płytki końcowej. Depolaryzacja rozprzestrzenia się następnie wzdłuż sarkolemmy, tworząc potencjał czynnościowy, ponieważ kanały sodowe przylegające do miejsca początkowej depolaryzacji wykrywają zmianę napięcia i otwierają się. Potencjał czynnościowy przemieszcza się po całej komórce, tworząc falę depolaryzacji.
ACh jest rozkładana przez enzym acetylocholinoesterazę (AChE) na acetyl i cholinę. AChE przebywa w szczelinie synaptycznej, rozkładając ACh tak, aby nie pozostawała związana z receptorami ACh, co spowodowałoby niepożądany przedłużony skurcz mięśnia (rysunek 6.9).
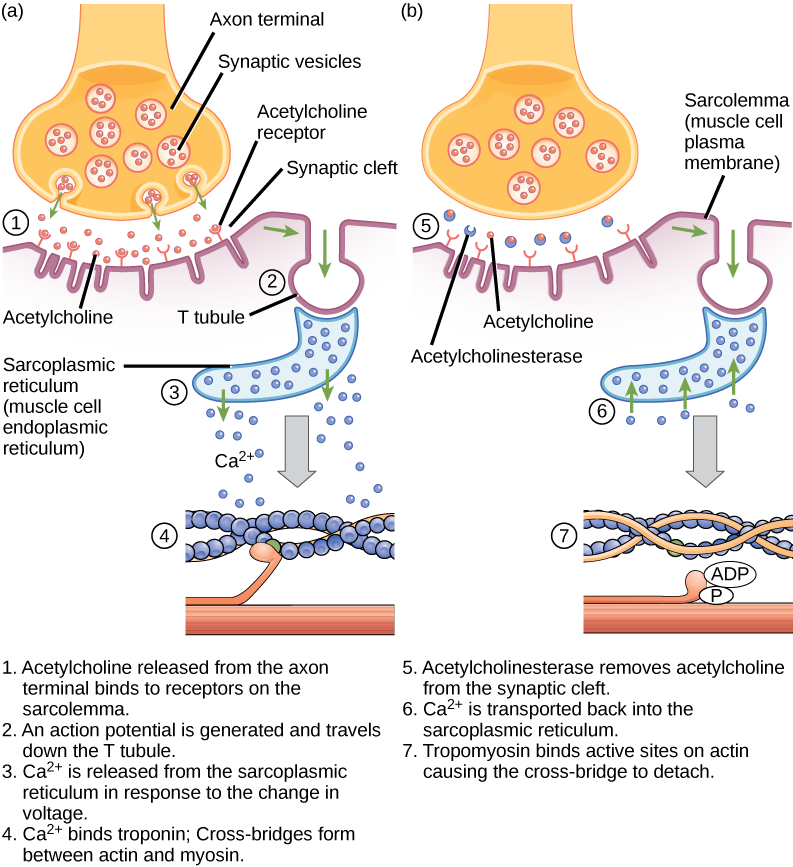
Rysunek 6.9. Ten schemat przedstawia sprzężenie pobudzenie-skurcz w skurczu mięśnia szkieletowego. Retikulum sarkoplazmatyczne jest wyspecjalizowanym retikulum endoplazmatycznym występującym w komórkach mięśniowych.
Ćwiczenia fizyczne
Śmiercionośny gaz nerwowy Sarin nieodwracalnie hamuje acetylocholinesterazę. Jaki wpływ miałby sarin na skurcz mięśnia?
Po depolaryzacji błona powraca do stanu spoczynku. Nazywa się to repolaryzacją, podczas której zamykają się kanały sodowe bramkowane napięciem. Kanały potasowe kontynuują przewodzenie na poziomie 90%. Ponieważ ATPaza sodowo-potasowa błony plazmatycznej zawsze transportuje jony, stan spoczynku (ujemnie naładowane wnętrze w stosunku do zewnątrz) zostaje przywrócony. Okres bezpośrednio po przekazaniu impulsu w nerwie lub mięśniu, w którym neuron lub komórka mięśniowa odzyskuje zdolność do przekazania kolejnego impulsu, nazywany jest okresem refrakcji. Podczas okresu refrakcji błona nie może wygenerować kolejnego potencjału czynnościowego. Okres refrakcji pozwala wrażliwym na napięcie kanałom jonowym na powrót do ich konfiguracji spoczynkowej. ATPaza sodowo-potasowa nieustannie przemieszcza Na+ z powrotem z komórki i K+ z powrotem do komórki, a K+ wycieka na zewnątrz, pozostawiając za sobą ładunek ujemny. Bardzo szybko błona repolaryzuje się, tak że może być ponownie zdepolaryzowana.
.