Egerület-összehúzódás kapcsolás
Az gerjesztés-összehúzódás kapcsolás a szarkolemmában keletkező akciós potenciál és az izomösszehúzódás megindulása közötti kapcsolat (transzdukció). A szarkoplazmatikus retikulumból a szarkoplazmába történő kalciumfelszabadulás kiváltó oka egy idegi jel. Minden egyes vázizomrostot egy-egy motoros neuron irányít, amely az agyból vagy a gerincvelőből az izomhoz továbbítja a jeleket. Az izomrost szarkolemmájának azt a területét, amely kölcsönhatásba lép a neuronnal, motoros véglemeznek nevezzük. A neuron axonjának végét szinaptikus terminálnak nevezik, és valójában nem érintkezik a motoros véglemezzel. A szinaptikus terminált és a motoros véglemezt egy kis tér, a szinaptikus hasadék választja el egymástól. Az elektromos jelek a neuron axonja mentén haladnak, amely az izomban elágazik, és a neuromuszkuláris csomópontban csatlakozik az egyes izomrostokhoz.
A sejtek elektromos kommunikációs képessége megköveteli, hogy a sejtek energiát fordítsanak arra, hogy elektromos gradienst hozzanak létre a sejtmembránjukon keresztül. Ezt a töltésgradienst az ionok szállítják, amelyek differenciáltan oszlanak el a membránon. Minden ion egy elektromos és egy koncentrációs hatást gyakorol. Ahogy a tej végül keveredés nélkül is elkeveredik a kávéval, úgy az ionok is egyenletesen oszlanak el, ha hagyjuk, hogy ez megtörténjen. Ebben az esetben nem engedik, hogy visszatérjenek az egyenletesen kevert állapotba.
A nátrium-kálium-ATPáz a sejt energiáját arra használja, hogy a K+ ionokat a sejt belsejében, a Na+ ionokat pedig a sejten kívül mozgatja. Ez önmagában kis elektromos töltést, de nagy koncentrációgradienst halmoz fel. A sejtben sok K+ van, a sejten kívül pedig sok Na+. A kálium az idő 90%-ában nyitott K+ csatornákon keresztül képes elhagyni a sejtet, és ezt meg is teszi. A Na+ csatornák azonban ritkán nyitva vannak, így a Na+ a sejten kívül marad. Amikor a K+ a koncentrációgradiensnek engedelmeskedve elhagyja a sejtet, az gyakorlatilag negatív töltést hagy maga után. Tehát nyugalmi állapotban nagy koncentrációs gradiens van a Na+ bejutására a sejtbe, és a sejtben hátrahagyott negatív töltések felhalmozódnak. Ez a nyugalmi membránpotenciál. A potenciál ebben az összefüggésben az elektromos töltés olyan elválasztását jelenti, amely képes munkát végezni. Voltban mérik, akárcsak egy akkumulátort. A transzmembránpotenciál azonban lényegesen kisebb (0,07 V); ezért a kis értéket millivoltban (mV) vagy 70 mV-ban fejezik ki. Mivel a sejt belseje negatív a külsőhöz képest, a mínusz jel a sejt belsejében lévő negatív töltések többletét, -70 mV-ot jelent.
Ha egy esemény megváltoztatja a membrán áteresztőképességét a Na+ ionok számára, akkor azok bejutnak a sejtbe. Ez megváltoztatja a feszültséget. Ez egy elektromos esemény, amelyet akciós potenciálnak nevezünk, és amely a sejt jeleként használható. Az idegek és az izmok között neurotranszmittereken keresztül történik a kommunikáció. Az idegi akciós potenciálok hatására neurotranszmitterek szabadulnak fel a szinaptikus terminálból a szinaptikus hasadékba, ahol aztán a szinaptikus hasadékon keresztül diffundálhatnak, és a motoros véglemezen lévő receptormolekulához kötődhetnek. A motoros véglemez a szarkolemmában olyan csomóponti redőkkel rendelkezik, amelyek nagy felületet biztosítanak a neurotranszmitter receptorokhoz való kötődéséhez. A receptorok valójában nátriumcsatornák, amelyek megnyílnak, hogy a neurotranszmitter jelének befogadásakor lehetővé tegyék a Na+ bejutását a sejtbe.
Az acetilkolin (ACh) a motoros neuronok által kibocsátott neurotranszmitter, amely a motoros véglemez receptoraihoz kötődik. A neurotranszmitter felszabadulása akkor következik be, amikor egy akciós potenciál lefelé halad a motoros neuron axonján, ami a szinaptikus terminális membrán megváltozott permeabilitását és kalcium beáramlását eredményezi. A Ca2+ ionok lehetővé teszik a szinaptikus vezikulák számára, hogy a preszinaptikus membránhoz (a neuronon) mozogjanak és kapcsolódjanak, és neurotranszmittert szabadítsanak fel a vezikulákból a szinaptikus hasadékba. Miután a szinaptikus terminálból felszabadult, az ACh a szinaptikus hasadékon keresztül a motoros véglemezre diffundál, ahol az ACh-receptorokhoz kötődik. Ahogy a neurotranszmitter kötődik, ezek az ioncsatornák megnyílnak, és a Na+ ionok átjutnak a membránon az izomsejtbe. Ez csökkenti a sejt belseje és külseje közötti feszültségkülönbséget, amit depolarizációnak nevezünk. Mivel az ACh a motoros véglemezhez kötődik, ezt a depolarizációt véglemezpotenciálnak nevezzük. A depolarizáció ezután a szarkolemma mentén terjed, és akciós potenciált hoz létre, mivel a kezdeti depolarizációs helyhez közeli nátriumcsatornák érzékelik a feszültségváltozást, és megnyílnak. Az akciós potenciál az egész sejten végighalad, és depolarizációs hullámot hoz létre.
Az acetilkolinészteráz (AChE) enzim az ACh-t acetilre és kolinra bontja. Az AChE a szinaptikus hasadékban tartózkodik, és úgy bontja le az ACh-t, hogy az ne maradjon az ACh-receptorokhoz kötődve, ami nem kívántan meghosszabbodott izomösszehúzódást okozna (6.9. ábra).
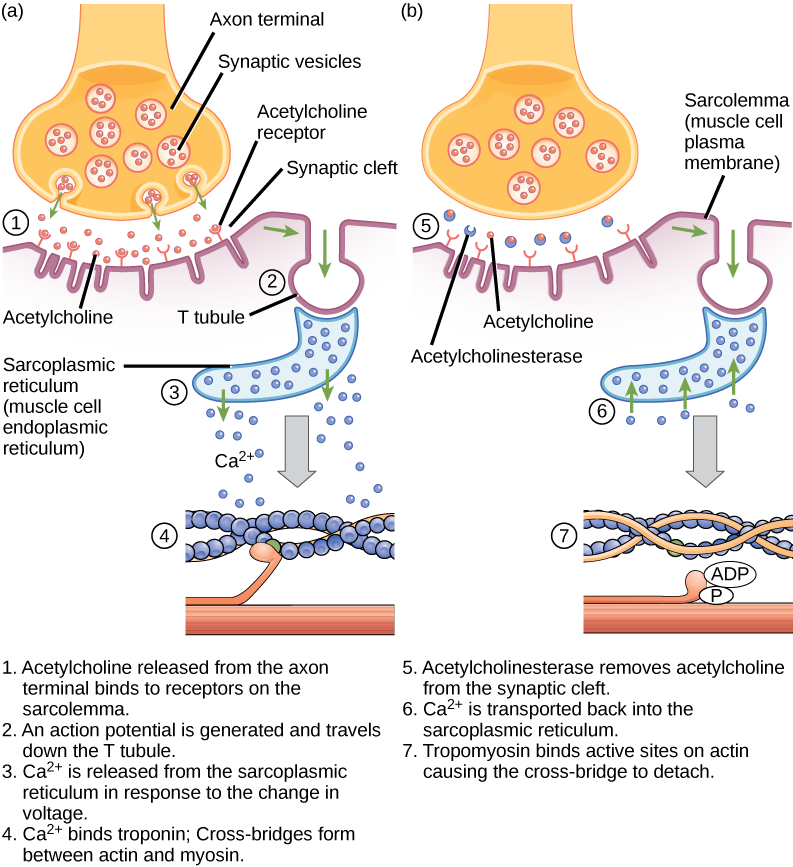
6.9. ábra. Ez az ábra a gerjesztés-összehúzódás kapcsolást mutatja a vázizomzat összehúzódásában. A szarkoplazmatikus retikulum az izomsejtekben található speciális endoplazmatikus retikulum.
Az edzés \(\PageIndex{1}\)
A halálos ideggáz szarin visszafordíthatatlanul gátolja az acetilkolinészterázt. Milyen hatással lenne a szarin az izomösszehúzódásra?
A depolarizáció után a membrán visszatér nyugalmi állapotába. Ezt nevezzük repolarizációnak, amely során a feszültségkapcsolt nátriumcsatornák bezáródnak. A káliumcsatornák továbbra is 90%-os vezetőképességgel működnek. Mivel a plazmamembrán nátrium-kálium ATPáza mindig ionokat szállít, a nyugalmi állapot (belül negatív töltésű a külsőhöz képest) visszaáll. Az idegben vagy izomban egy impulzus továbbítását közvetlenül követő időszakot, amelyben az ideg- vagy izomsejt visszanyeri képességét egy újabb impulzus továbbítására, refrakter periódusnak nevezzük. A refrakter időszak alatt a membrán nem képes újabb akciós potenciált létrehozni. A refrakter időszak lehetővé teszi a feszültségérzékeny ioncsatornák számára, hogy visszatérjenek nyugalmi konfigurációjukba. A nátrium-kálium-ATPáz folyamatosan mozgatja vissza a Na+-ot a sejtből és a K+-t a sejtbe, a K+ pedig negatív töltést hátrahagyva szivárog ki. A membrán nagyon gyorsan repolarizálódik, így ismét depolarizálódhat.