Excitation-contraction coupling
Excitation-contraction coupling on yhteys (transduktio) sarkolemmassa syntyvän toimintapotentiaalin ja lihassupistuksen alkamisen välillä. Laukaiseva tekijä kalsiumin vapautumiselle sarkoplasmisesta retikulumista sarkoplasmaan on hermosignaali. Jokaista luurankolihaskuitua ohjaa motoneuroni, joka välittää signaaleja aivoista tai selkäytimestä lihakseen. Lihassyyn sarkolemman aluetta, joka on vuorovaikutuksessa hermosolun kanssa, kutsutaan motoriseksi päätylevyksi. Neuronin aksonin päätä kutsutaan synaptiseksi terminaaliksi, eikä se itse asiassa ole kosketuksissa motoriseen päätylevyyn. Synaptinen pääte ja motorinen päätelevy ovat erillään pienessä tilassa, jota kutsutaan synaptiseksi rakoiluksi. Sähköiset signaalit kulkevat neuronin aksonia pitkin, joka haarautuu lihaksen läpi ja yhdistyy yksittäisiin lihassäikeisiin neuromuskulaarisessa liitoskohdassa.
Solujen kyky kommunikoida sähköisesti edellyttää, että solut kuluttavat energiaa luodakseen sähköisen gradientin solukalvojensa yli. Tätä varausgradienttia kuljettavat ionit, jotka jakautuvat eri tavoin kalvon poikki. Kullakin ionilla on sekä sähköinen että konsentraatiovaikutus. Aivan kuten maito lopulta sekoittuu kahviin ilman sekoittamista, myös ionit jakautuvat tasaisesti, jos niiden annetaan tehdä niin. Tällöin niiden ei anneta palata tasaisesti sekoittuneeseen tilaan.
Natrium-kalium-ATPaasi käyttää soluenergiaa K+-ionien siirtämiseen solun sisällä ja Na+-ionien siirtämiseen solun ulkopuolella. Tämä yksinään kerryttää pienen sähkövarauksen, mutta suuren konsentraatiogradientin. Solun sisällä on paljon K+ ja solun ulkopuolella paljon Na+. Kalium pystyy poistumaan solusta K+-kanavien kautta, jotka ovat auki 90 % ajasta, ja niin se tekeekin. Na+ -kanavat ovat kuitenkin harvoin auki, joten Na+ jää solun ulkopuolelle. Kun K+ poistuu solusta konsentraatiogradienttiaan noudattaen, se jättää jälkeensä negatiivisen varauksen. Lepotilassa Na+:lle on siis suuri konsentraatiogradientti solun sisälle, ja soluun jäävät negatiiviset varaukset kasautuvat. Tämä on kalvon lepopotentiaali. Potentiaali tarkoittaa tässä yhteydessä sähkövarauksen erottumista, joka pystyy tekemään työtä. Se mitataan voltteina, aivan kuten paristossa. Transmembraanipotentiaali on kuitenkin huomattavasti pienempi (0,07 V); siksi pieni arvo ilmaistaan millivolteina (mV) eli 70 mV:nä. Koska solun sisäpuoli on negatiivinen verrattuna ulkopuoliin, miinusmerkki tarkoittaa negatiivisten varausten ylimäärää solun sisällä, -70 mV.
Jos jokin tapahtuma muuttaa kalvon läpäisevyyttä Na+ -ioneille, ne pääsevät soluun. Se muuttaa jännitettä. Tämä on sähköinen tapahtuma, jota kutsutaan toimintapotentiaaliksi ja jota voidaan käyttää solun signaalina. Hermojen ja lihasten välillä tapahtuu viestintää välittäjäaineiden välityksellä. Hermoston toimintapotentiaalit aiheuttavat välittäjäaineiden vapautumisen synaptisesta päätepisteestä synaptiseen rakoon, josta ne voivat sitten diffundoitua synaptisen rakon läpi ja sitoutua motorisen päätelevyn reseptorimolekyyliin. Motorisella päätylevyllä on sarkolemmassa liitospoimuja – poimuja, jotka luovat suuren pinta-alan välittäjäaineen sitoutumiselle reseptoreihin. Reseptorit ovat itse asiassa natriumkanavia, jotka avautuvat päästääkseen Na+:n kulkemaan soluun, kun ne vastaanottavat välittäjäainesignaalin.
Asetyylikoliini (ACh) on motoneuronien vapauttama välittäjäaine, joka sitoutuu motorisen päätylevyn reseptoreihin. Neurotransmitterin vapautuminen tapahtuu, kun toimintapotentiaali kulkee motoneuronin aksonia pitkin, jolloin synaptisen päätelaitteen kalvon läpäisevyys muuttuu ja kalsiumia virtaa sisään. Ca2+ -ionien ansiosta synaptiset vesikkelit voivat siirtyä presynaptiselle kalvolle (neuronissa) ja sitoutua siihen sekä vapauttaa välittäjäainetta vesikkeleistä synaptiseen rakoon. Kun ACh on vapautunut synaptisesta päätepisteestä, se diffundoituu synaptisen raon läpi motoriseen päätelevyyn, jossa se sitoutuu ACh-reseptoreihin. Kun välittäjäaine sitoutuu, nämä ionikanavat avautuvat, ja Na+ -ionit ylittävät kalvon lihassoluun. Tämä pienentää solun sisä- ja ulkopinnan välistä jännite-eroa, mitä kutsutaan depolarisaatioksi. Koska ACh sitoutuu motoriseen päätelevyyn, tätä depolarisaatiota kutsutaan päätelevypotentiaaliksi. Tämän jälkeen depolarisaatio leviää pitkin sarkolemmaa luoden toimintapotentiaalin, kun alkuperäisen depolarisaatiokohdan vieressä olevat natriumkanavat havaitsevat jännitteen muutoksen ja avautuvat. Toimintapotentiaali siirtyy koko solun läpi, jolloin syntyy depolarisaatioaalto.
Acetyylikoliiniesteraasi (AChE) -entsyymi hajottaa ACh:n asetyyliksi ja koliiniksi. AChE sijaitsee synaptisessa raossa ja hajottaa ACh:ta, jotta se ei jäisi sitoutumaan ACh-reseptoreihin, mikä aiheuttaisi ei-toivottua pitkittynyttä lihassupistusta (kuva 6.9).
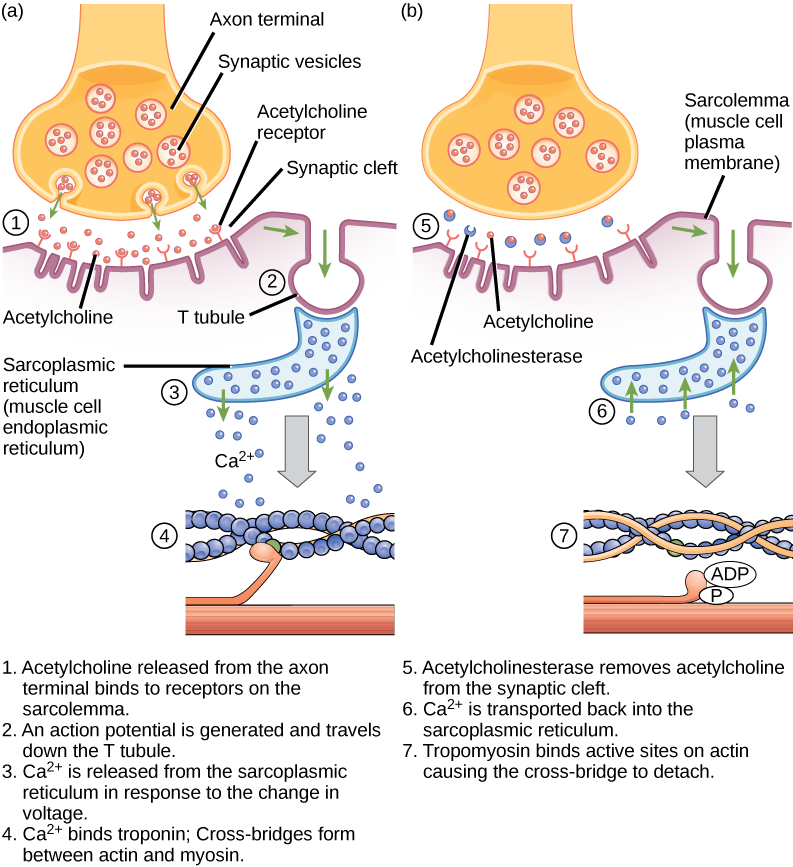
Kuva 6.9. Tässä kaaviossa on esitetty heräte-kontraktiokytkentä luurankolihaksen supistumisessa. Sarkoplasminen retikulum on lihassoluissa esiintyvä erikoistunut endoplasminen retikulum.
Harjoitus \(\PageIndex{1}\)
Tappava hermokaasu Sarin estää peruuttamattomasti asetyylikoliiniesteraasia. Mikä vaikutus Sarinilla olisi lihassupistukseen?
Depolarisaation jälkeen kalvo palaa lepotilaan. Tätä kutsutaan repolarisaatioksi, jonka aikana jänniteohjatut natriumkanavat sulkeutuvat. Kaliumkanavat jatkavat 90 %:n johtavuudella. Koska plasmakalvon natrium-kalium-ATPaasi kuljettaa aina ioneja, lepotila (negatiivisesti varautunut sisäpuoli ulkoilmaan nähden) palautuu. Välittömästi hermo- tai lihasimpulssin välittämistä seuraavaa ajanjaksoa, jolloin hermosolu tai lihassolu palauttaa kykynsä välittää uutta impulssia, kutsutaan refraktoriseksi ajanjaksoksi. Refraktorisen jakson aikana kalvo ei voi synnyttää uutta toimintapotentiaalia. Refraktorijakso antaa jänniteherkille ionikanaville mahdollisuuden palata lepokokoonpanoonsa. Natrium-kalium-ATPaasi siirtää jatkuvasti Na+ takaisin ulos solusta ja K+ takaisin soluun, ja K+ vuotaa ulos jättäen jälkeensä negatiivisen varauksen. Hyvin nopeasti kalvo repolarisoituu, jotta se voi jälleen depolarisoitua.