Úvod
Lidský mozek prochází během života rozsáhlým zráním, které usnadňuje kognitivní vývoj. Jedním z takových zásadních procesů zrání je myelinizace axonů v celém nervovém systému. V centrální nervové soustavě (CNS) rozšiřují gliové buňky zvané oligodendrocyty do svého okolí mnoho výběžků, které koncentricky obalují axony membránou a vytvářejí tak myelinové pochvy. Myelinové pochvy umožňují rychlé solární vedení akčních potenciálů tím, že lokalizují napěťově řízené Na+ kanály do krátkých mezer mezi sousedními pochvami (tzv. Ranvierovy uzly) a působí jako elektrické izolátory. Axony, které jsou plně myelinizované po celé své délce, vedou impulzy mnohonásobně rychleji než nemyelinizované axony o stejné velikosti průřezu (Waxman, 1980). Proto myelinizované nervové obvody vedou informace mnohem rychleji než obvody nemyelinizované. Lidé se rodí s prakticky nemyelinizovaným CNS a populace oligodendrocytů se po narození dramaticky rozšíří a v prvních letech dětství dochází k rozsáhlé myelinizaci. Myelinizace pokračuje během dospívání a v dospělosti charakteristickým časoprostorovým způsobem, což koreluje se vznikem a udržováním správné funkce obvodů. Například dozrávání bílé hmoty (oblastí CNS bohatých na myelin) probíhá současně s vývojem kognitivních procesů v dětství, jako je rychlost zpracování informací (Mabbott et al., 2006; Scantlebury et al., 2014). Kromě toho se patologie/abnormality myelinu vyskytují nejen u demyelinizačního onemocnění roztroušená skleróza, ale také u několika neurodegenerativních onemocnění (Kang et al., 2013; Huang et al., 2015) a neurovývojových poruch (Takahashi et al., 2011). Myelinizace jednotlivých axonů však není jevem „všechno nebo nic“. Axony v CNS vykazují rozsáhlé rozdíly v počtu myelinových pochev, jejich délce, tloušťce a rozložení po celé délce. Existuje mnoho různých vzorů myelinizace; například axony s řídce myelinizovanými oblastmi byly popsány v juvenilní i dospělé myší kůře (Tomassy et al., 2014; Hill et al., 2018; Hughes et al., 2018). Modifikace kteréhokoli z těchto parametrů pláště má předvídatelné účinky na rychlost vedení podkladového axonu – proto může být stanovení specifického vzoru myelinizace podél axonu obzvláště důležité pro vyladění funkce obvodu. Například axony ve sluchovém mozkovém kmeni pískomilů vykazují podél distálních oblastí postupně kratší myelinové pochvy, které zajišťují přesné načasování příchodu signálu a usnadňují lokalizaci zvuku (Ford et al., 2015). Jemné změny v celkovém vzoru myelinizace podél axonu (buď přidáním nového myelinu, nebo remodelací stávajícího myelinu) by mohly zásadně změnit načasování nervových impulzů v obvodech. Pokud je myelin adaptabilní, pak může modifikace takových vzorců myelinizace představovat mocný mechanismus regulace funkce obvodů v průběhu života.
Nejnovější důkazy naznačují, že myelin může být adaptabilní v reakci na aktivitu obvodů. Difuzní tenzorové zobrazování celého mozku lze použít k měření rozsáhlých změn v bílé hmotě bohaté na myelin v průběhu času – takové experimenty u lidí a hlodavců ukázaly, že učení se novému úkolu koreluje se změnami bílé hmoty v příslušných oblastech mozku (Scholz et al., 2009; Sampaio-Baptista et al., 2013). Analýzy na buněčné úrovni na zvířecích modelech ukazují, že pro efektivní motorické učení je nutná produkce nových myelinizačních oligodendrocytů (McKenzie et al., 2014; Xiao et al., 2016). V současné době se předpokládá, že aktivita neuronálního okruhu může vyvolat změny v myelinu; rozsáhlý výzkum prokázal, že neuronální aktivita může ovlivnit proliferaci prekurzorových buněk oligodendrocytů (OPC), diferenciaci oligodendrocytů a tvorbu a růst myelinových pochev. Tento výzkum, včetně důkazů o příslušných molekulárních signálech, byl rozsáhle rozebrán jinde (Fields, 2015; Almeida a Lyons, 2017; Mount a Monje, 2017). Neuronální aktivita by mohla pohánět změny myelinu, které by zase mohly měnit rychlost vedení a dolaďovat tak časování, jež je základem funkce obvodů.
Stále však nevíme, zda a jak je myelinizace obvodů dynamicky regulována v průběhu života. Práce na hlodavcích naznačují, že nové oligodendrocyty vznikají v celém CNS i v dospělosti (Young et al., 2013), a OPC se skutečně nacházejí v lidském dospělém mozku (Chang et al., 2000). Uhlíková analýza lidské tkáně identifikovala v kůře oligodendrocyty narozené v dospělosti, ačkoli stejné analýzy ukázaly, že většina oligodendrocytů v corpus callosum vzniká v raném dětství (Yeung et al., 2014). Neurozobrazovací studie u lidí, které korelují strukturální změny bílé hmoty s učením úkolů, však naznačují, že nový myelin se může tvořit po celý život. Takto prolongovaná myelinizace by v zásadě vyžadovala celoživotní tvorbu oligodendrocytů, vzhledem k tomu, že jednotlivé myelinizující oligodendrocyty mají omezené časové okno pouhých několika hodin k zahájení tvorby nových pochev (Watkins et al., 2008; Czopka et al., 2013) a počet pochev na oligodendrocyt se zdá být v čase stabilní (Tripathi et al., 2017). Jedna výhrada, na kterou upozorňují Mount a Monje (2017), spočívá v tom, že datum „narození“ v experimentu s datováním uhlíku (které určuje časový okamžik replikace DNA během buněčného dělení), odráží datum OPC, nikoli nutně diferencovaného oligodendrocytu. To je důležité vzhledem k důkazům, že OPC se mohou přímo diferencovat v oligodendrocyty bez buněčného dělení, přinejmenším u hlodavců (Hughes et al., 2013). OPC v corpus callosum se mohly přímo diferencovat v oligodendrocyty mnoho let po jejich konečném buněčném dělení; dobu diferenciace těchto nových oligodendrocytů tedy nelze určit pomocí datování uhlíku, a proto Yeung et al. (2014) možná podcenili rychlost produkce oligodendrocytů v dospělém lidském mozku. Stále se máme co učit o relativním podílu tvorby oligodendrocytů a remodelace myelinu na vývoji CNS v průběhu života.
Pro úplné pochopení přesné dynamiky oligodendrogeneze, tvorby myelinu a remodelace myelinu v různých fázích života představuje longitudinální zobrazování s vysokým rozlišením zlatý standard. Zde uvádíme přehled nedávných zobrazovacích studií in vivo, které začínají objasňovat dynamiku myelinizace, což nám také umožní začít chápat, jak tato dynamika může ovlivňovat funkci nervových obvodů.
De novo myelinizace
Aby bylo možné začít definitivně řešit, jak vznikají oligodendrocyty a jak se tvoří a dynamicky remodeluje myelin in vivo, byly ve dvou nedávných studiích použity opakované dvoufotonové snímky somatosenzorické kůry myší po delší dobu. Hughes et al. (2018) zobrazovali kůru transgenních reportérových myší s fluorescenčně značenými buňkami oligodendrogliální linie od rané dospělosti, přes střední a starší věk (přibližně P720). Zjistili, že populace oligodendrocytů se nadále rozšiřuje a že hustota kortikálních oligodendrocytů se mezi stadii mladého dospělého a středního věku téměř zdvojnásobuje (obr. 1A). To bylo doprovázeno více než dvojnásobným nárůstem počtu kortikálních myelinových pochev. Jak se ale počet oligodendrocytů zvyšuje? V časném postnatálním vývoji vzniká mnoho oligodendrocytů, ale pouze podskupina přežije a pokračuje v myelinizaci axonů (Barres et al., 1992). Zdá se, že podobně je tomu i v dospělosti – sledováním jednotlivých kortikálních OPC v dospělé kůře po dobu až 50 dní Hughes et al. (2018) odhalili, že většina nově diferencovaných oligodendrocytů podléhá buněčné smrti a pouze 22 % jich přežije a zapojí se do myelinizace (obr. 1B). Stále není známo, jaký podíl nově diferencovaných oligodendrocytů vzniká po dělení OPC oproti přímé diferenciaci. Jakmile se však oligodendrocyty zaváží k myelinizaci, zůstávají stabilní, přičemž během 50denního zobrazovacího období nebyly nalezeny žádné důkazy o tom, že by myelinizující oligodendrocyty podléhaly buněčné smrti.
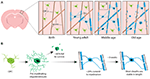
OBRÁZEK 1. Dynamika oligodendrocytů a myelinu v mozkové kůře savců v průběhu života. (A) Oligodendrocytární prekurzorové buňky (OPC) kontinuálně vytvářejí nové myelinizující oligodendrocyty (OL) v somatosenzorické kůře od narození až do středního věku. Ve stáří pak populace OL klesá, což je doprovázeno snížením myelinového pokrytí. (B) Lineární sledování jednotlivých OPC ukazuje, že ačkoli premyelinizační OL vznikají v dospělosti nepřetržitě, pouze přibližně 20 % z nich přežije, aby myelinizovaly. Většina jednou vytvořených myelinových pochev je po delší dobu stabilní v délce, což naznačuje, že za normálních okolností dochází k velmi malé remodelaci stávajícího myelinu. Shrnutí údajů od Hill et al. (2018) a Hughes et al. (2018):
Podobně Hill et al. (2018) použili transgenní reportéry oligodendrocytů a techniku bezznačkové spektrální konfokální reflektance (SCoRe) mikroskopie k zobrazení myelinu podél axonů (Schain et al., 2014) v somatosenzorické kůře juvenilních, mladých dospělých, středně starých a starých myší (P950). Zjistili také, že počet oligodendrocytů se v dospělosti dále zvyšuje až do P650 a že oligodendrocyty jsou ve středním věku stabilní až do 80 dnů zobrazování. Zjistili, že myelinizace kůry dosahuje vrcholu také ve středním věku v P650 a že hustota oligodendrocytů od svého vrcholu (v P650) až do velmi vysokého věku (P950) výrazně klesá (obrázek 1A). To se projevilo snížením myelinového pokrytí korových axonů vrstvy I mezi P650 a P950. Dlouhodobé přežívání oligodendrocytů se může v různých částech CNS lišit. Tripathi et al. (2017) označili myší myelinizační oligodendrocyty v P60 a poté spočítali, kolik označených buněk přežilo do P605 v několika oblastech CNS. Zjistili, že v míše a motorické kůře přežilo 60-70 % buněk označených P60, zatímco v corpus callosum přežilo více než 90 % buněk označených P60. Snižování počtu oligodendrocytů a myelinizace v některých oblastech CNS s věkem vyvolává zajímavé otázky týkající se úlohy úbytku myelinu při zhoršování kognitivních funkcí souvisejících s věkem. Analýza MRI ukazuje, že mikrostruktura bílé hmoty koreluje s fluidní inteligencí (Ritchie et al., 2015), ale také že se tato mikrostruktura bílé hmoty s rostoucím věkem zhoršuje (Cox et al., 2016). Následná ztráta myelinu spojená s věkem by mohla vést ke snížení kognitivních funkcí v důsledku dysregulace myelinových okruhů.
Mohla by tvorba nových oligodendrocytů (a následně nového myelinu) v kůře dospělých reagovat na aktivitu okruhů? Předchozí výzkum ukázal, že snížení senzorického vstupu odstraněním fousků u myší vede ke snížení oligodendrogeneze v somatosenzorické kůře (Hill et al., 2014). Za účelem dalšího zkoumání této skutečnosti poskytovali Hughes et al. (2018) dospělým myším (P365) po dobu 3 týdnů senzorickou stimulaci zavěšením korálků do zvířecích klecí, které opakovaně stimulovaly jejich vousky, a tím i somatosenzorickou kůru. Zobrazením somatosenzorické kůry před a po 3 týdnech prokázali, že senzorická stimulace zvyšuje počet oligodendrocytů, což je potenciálně způsobeno zvýšeným přežíváním nově diferencovaných buněk. Kougioumtzidou et al. (2017) poskytli další důkaz, že aktivita obvodů může být důležitá pro regulaci přežívání buněk – prokázali, že ztráta podjednotek AMPA receptorů 2, 3 a 4 v OPC vede ke sníženému přežívání oligodendrocytů. To naznačuje, že myelinizace de novo by mohla být modulována aktivitou kortikálních okruhů v průběhu života, možná za účelem doladění funkce týchž okruhů.
Zůstává mnoho otázek, které je třeba vyřešit: jaký je vliv oligodendrogeneze a nové myelinizace na skutečnou funkci okruhů? Zvyšuje neuronální aktivita dlouhodobé přežívání myelinizujících oligodendrocytů? Je možné, že úbytek oligodendrocytů ve stáří je způsoben snížením neuronální aktivity souvisejícím s věkem, což by zase mohlo ovlivnit celkové přežívání oligodendrocytů. Případně je možné, že oligodendrocyty mají omezenou životnost nezávisle na aktivitě neuronů (buď vnitřně naprogramovanou, nebo ovlivněnou jinými vnějšími signály spojenými se stárnutím). V obou případech by stimulace obvodů mohla pomoci zmírnit úbytek myelinu související s věkem buď podporou přežívání stávajících oligodendrocytů, nebo stimulací tvorby nových oligodendrocytů. To by zase mohlo mít významné důsledky pro léčbu a prevenci poklesu kognitivních funkcí souvisejících s věkem.
Aktivitou zprostředkovaná oligodendrogeneze se neomezuje pouze na somatosenzorickou kůru – mladé dospělé myši podstupující motorické učení rovněž vykazují zvýšení počtu nově diferencovaných oligodendrocytů v motorické kůře (Xiao et al., 2016). A co ostatní oblasti CNS? Mnoho kortikálních axonů se promítá přes corpus callosum, a proto by stimulace kortikálních okruhů mohla signalizovat jak kortikální, tak kalózní OPC. Dvě studie na hlodavcích prokázaly, že stimulace kortikálních neuronů indukuje oligodendrogenezi v corpus callosum. Gibson et al (2014) optogeneticky stimulovali projekční neurony vrstvy V v premotorické kůře a zjistili nárůst proliferace OPC jak v premotorické kůře, tak v corpus callosum. To vedlo ke zvýšení počtu oligodendrocytů a tloušťky pochev 4 týdny po stimulaci. Nedávno Mitew et al. (2018) použili Designer Receptors Exclusively Activated by Designer Drugs ke stimulaci somatosenzorických neuronů vrstvy 2/3 a rovněž pozorovali zvýšenou proliferaci OPC, oligodendrogenezi a silnější myelinové pochvy v corpus callosum u mladých i dospělých myší. Prokázali také, že nové oligodendrocyty přednostně tvoří myelinové pochvy na aktivních axonech. To naznačuje, že myelinizace de novo vyvolaná aktivitou může být v zásadě zaměřena na aktivní axony/obvody. Zatím není známo, jak dlouhodobé mohou být změny myelinu v reakci na neuronální aktivitu. Dlouhodobé přežívání myelinizačních buněk, které zaznamenali Tripathi et al. (2017), Hill et al. (2018) a Hughes et al. (2018), naznačuje, že jakmile oligodendrocyt vytvoří myelinové pochvy, pravděpodobně přežije, i když se úroveň neuronální aktivity vrátí zpět na výchozí úroveň. To, zda se samotné myelinové pochvy změní, jakmile se neuronální aktivita vrátí na normální úroveň, vyžaduje další zkoumání dynamiky jednotlivých pochev, které je popsáno níže.
Je tedy možné, že v mnoha oblastech CNS, kde mají axony vhodné pro myelinizaci dostatek nemyelinizovaného prostoru, může docházet k celoživotní de novo myelinizaci. Zůstává však nejasné, do jaké míry oligodendrogeneze pokračuje v různých oblastech dospělého lidského mozku. Analýza datování uhlíku naznačuje, že většina oligodendrocytů v traktu corpus callosum vzniká v raném dětství (Yeung et al., 2014). Imunohistochemická analýza lidské mozkové tkáně pomocí nového markeru pro nově diferencované oligodendrocyty (BCAS1) ukazuje nové oligodendrocyty ve frontální kůře i po středním věku, ale velmi málo nových oligodendrocytů v bílé hmotě po třetí dekádě života (Fard et al., 2017). Tento rozdíl v oligodendrogenezi mezi jednotlivými druhy může být důsledkem rozsahu. Údaje Hughese et al. z výzkumu hlodavců (2013) naznačují, že oligodendrocytů vzniká obrovský nadbytek, přičemž průběžně dochází k prořezávání téměř 80 % buněk. Vzhledem k energetickým nákladům takového procesu, je tento mechanismus udržitelný po celý život v orgánu velikosti lidského mozku? Možná, že v lidském mozku dochází k omezené nadprodukci oligodendrocytů z důvodu potřeby delší myelinizace většího CNS nebo proto, že signály, jako je neuronální aktivita, stimulují OPC k diferenciaci na oligodendrocyty podle potřeby.
Přeměna myelinu
Přeměna stávajících myelinových pochev by mohla změnit vlastnosti vedení bez nutnosti generovat nové oligodendrocyty nebo myelin. Změna délky stávajících myelinových pochev by mohla změnit pokrytí myelinem podél axonu a vzdálenost mezi Ranvierovými uzly (což by obojí ovlivnilo rychlost vedení). Kromě toho by i velmi jemná remodelace myelinu mohla změnit délku samotných uzlů. Nedávno se ukázalo, že délka uzlů se může v optickém nervu a v kůře značně lišit a že změna délky uzlů podél axonu může v zásadě také významně změnit rychlost vedení (Arancibia-Cárcamo et al., 2017). Zda jsou změny Ranvierových uzlů primárně způsobeny myelinizací, nebo reorganizací samotného axonu, je třeba ještě určit.
Hill et al (2018) i Hughes et al (2018) provedli longitudinální studii jednotlivých myelinových pochev v somatosenzorické kůře myší po dobu několika týdnů, aby posoudili, zda je délka pochev dynamicky regulována. Hill et al (2018) zjistili, že v raném dospělém věku (P90-120) sice některé pochvy vykazují prodloužení nebo smrštění, ale 81 % pozorovaných pochv je stabilních. S věkem může být stabilní délka většího počtu pochev; Hughes et al. (2018) sledovali délku pochev u starších zvířat (P365) a zjistili, že 99 % pochev zůstalo stabilních po dobu 3 týdnů (obr. 1B).
Podobná stabilita délky pochev byla popsána i jinde; Auer et al. (2018) použili larvální zebřičky ke zkoumání, zda se délka jednotlivých pochev může měnit v čase, a to pomocí časosběrného živého zobrazování fluorescenčně značených myelinových pochev. Zjistili, že jednotlivé pochvy procházejí v prvních dnech po vzniku rychlým, ale proměnlivým růstem, než se jejich délka ustálí. Jakmile se stabilizují, pokračují v růstu pouze tak, aby se přizpůsobily celkovému růstu zvířete.
Proč se délka některých pochev v kůře mění, zatímco jiných ne? Může to odrážet různorodost požadavků odlišných nervových obvodů. Axonální diverzita byla pozorována během počáteční myelinizace v míše zebřiček, kde některé axony využívají uvolňování synaptických vezikul k regulaci počtu a délky myelinových pochev, zatímco jiné nikoli (Koudelka et al., 2016). To vyvolává zajímavou hypotézu, že pouze některé axony jsou schopny regulovat myelin prostřednictvím signálů souvisejících s aktivitou. Hughes et al. (2018) zjistili, že jejich paradigma senzorické stimulace nezvýšilo podíl dynamických pochev v somatosenzorické kůře. Podrobnější analýza diverzity podtypů axonů spojená s longitudinální studií dynamiky délky pochev by však mohla potvrdit, zda je remodelace délky pochev specifická pro určité okruhy.
Do sheath length stability reflect an inability of sheaths to remodel? Experimenty na zebřičkách naznačují, že remodelace délky pochvy může být vyvolána, když je narušen profil myelinizace axonu. Auer et al. (2018) ablovali jednotlivé oligodendrocyty, a proto řídce odstraňovali pochvy podél axonů. Zjistili, že při ztrátě jediné myelinové pochvy na plně myelinizovaném axonu mohou sousední pochvy znovu zahájit rychlý růst, aby pokryly nemyelinizovanou mezeru. V několika případech se nová myelinová pochva vytvořila na místě své předchůdkyně a mohla se dokonce tlačit zpět proti napadeným sousedním pochvám, aby obnovila původní vzorec myelinizace (obr. 2A). Někdy se tedy specifický vzorec myelinizace přednostně zachovává i po přerušení myelinu. Důvodem může být udržení optimalizovaných vodivostních vlastností základního axonu. Auer et al. (2018) pozorovali u larválních zebřiček řídce myelinizované axony, které byly dříve identifikovány v kůře hlodavců. Zajímavé je, že zjistili, že po ablaci jednotlivých pochev na takto řídce myelinizovaných axonech se vytvořila nová pochva prakticky na stejném místě jako ablovaná pochva, a to i podél jinak nemyelinizovaného úseku axonu (obr. 2B). Zdá se tedy, že vzorce myelinizace podél řídce myelinizovaných axonů se u zebřiček také stabilně udržují, jak naznačují Hill et al. (2018) u hlodavců. Funkce profilů řídké myelinizace zůstává neznámá. Takové vzory mohou umožnit dynamičtější vyladění funkce jednotlivých axonů v průběhu času, i když je také možné, že takové nemyelinizované mezery mohou usnadňovat postupnou myelinizaci pro zachování konzistentních dob vedení v rámci obvodů, jak zvíře roste a/nebo se mění délka axonů.
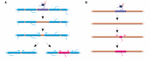
FIGURE 2. K remodelaci myelinu může docházet in vivo. (A) Ablace jednotlivých pochev na plně myelinizovaném axonu může vyvolat rychlý růst sousedních pochev, které mezeru překryjí. Tato mezera může být buď zcela pokryta sousedními pochvami, nebo může být přidáním nové pochvy obnoven původní profil myelinizace. (B) Po ablaci pochvy na řídce myelinizovaném axonu následuje tvorba nové myelinové pochvy identické velikosti a umístění jako ablovaná předchozí pochva. Shrnutí údajů z Auer et al. (2018).
Mají stabilní myelinové pochvy u savců také tuto schopnost remodelovat, když je narušen vzorec myelinizace? K zodpovězení této otázky jsou nutné další longitudinální studie spojené s demyelinizací. Je možné, že taková remodelace není vyvolána neuronální aktivitou, ale je kompenzačním mechanismem ztráty myelinu. Úbytek oligodendrocytů spojený s věkem by mohl vyvolat remodelaci přežívajících pochev, aby pokryly denudované části axonu, a tím pomohly zachovat funkci obvodu. Zda tomu tak skutečně je, by mohlo určit živé zobrazování myelinových pochev ve stáří.
Všechny zde diskutované studie živého zobrazování hodnotily dynamiku délky myelinových pochev, nikoliv však tloušťku myelinových pochev. Může být tloušťka pochev dynamicky regulována? Stimulace signalizace PI3K/AKT/mTOR v oligodendrocytech dospělých myší vyvolává dodatečné obalování myelinu, které zvyšuje tloušťku pochvy (Snaidero et al., 2014). To může být modulováno aktivitou obvodů, protože neuronální stimulace vede ke zvýšení tloušťky pochvy u juvenilních i dospělých myší (Gibson et al., 2014; Mitew et al., 2018). To zdůrazňuje potřebu zobrazovat všechny parametry pochvy longitudinálně, abychom plně porozuměli dynamice remodelace pochvy. K přesnému měření tloušťky pochvy podél axonů jsou zapotřebí zobrazovací metody v přímém přenosu, protože v současné době to vyžaduje měření v příčném řezu pomocí elektronové mikroskopie, což omezuje analýzu na jeden časový bod. Některé zobrazovací techniky bez použití značek, jako je mikroskopie třetí harmonické generace a spektrální reflektometrie, jsou pro provádění takových měření slibné (Lim et al., 2014; Kwon et al., 2017). Spojení těchto technik s longitudinálními studiemi kůry hlodavců by mohlo určit, zda zavedené myelinové pochvy mohou upravovat svou tloušťku, nebo zda neuronální aktivita jednoduše tlačí na de novo myelinizaci, která vytváří silnější pochvy.
Zdá se tedy, že ačkoli jsou myelinové pochvy schopny remodelace při narušení myelinu, většina pochev je obecně stabilní co do délky. Tato stabilita je potenciálně způsobena udržováním časně zavedených vzorců myelinizace optimalizovaných pro funkci obvodu.
Budoucnost
Nejnovější zobrazovací studie savců se zaměřily na de novo myelinizaci a remodelaci pochev v šedé kůře mozkové. Kortikální okruhy přijímají a vysílají informace prostřednictvím mnoha oblastí, jako je mícha a corpus callosum, a tak by změny myelinu v několika různých oblastech CNS mohly změnit signalizaci v jednom okruhu. CNS se tradičně popisuje podle vzhledu po fixaci formaldehydem, kde „bílá hmota“ popisuje silně myelinizované axonální dráhy, zatímco „šedá hmota“ popisuje oblasti hustě zaplněné těly neuronálních buněk, dendrity a synapsemi. Tato klasifikace je však příliš zjednodušující; OPC produkují myelinizační oligodendrocyty jak v šedé, tak v bílé hmotě (Dawson et al., 2003) a ve skutečnosti se objevují důkazy o rozmanitosti v oligodendrogliové linii a ve vzorcích myelinizace jak v šedé, tak v bílé hmotě (Rivers et al., 2008; Viganò et al., 2013; Young et al., 2013; Bechler et al., 2015). Tato rozmanitost může odrážet jedinečné požadavky na myelin v různých oblastech a potenciálně i na různé okruhy CNS. K lepšímu pochopení dynamiky de novo myelinizace a remodelace pochev v oblastech CNS mimo kůru jsou nutné další longitudinální zobrazovací studie.
Zatímco optická průhlednost larvální zebřičky umožňuje neinvazivní živé zobrazování, provádění takových experimentů v CNS savců je invazivnější a technicky náročnější. Hill et al. (2018) a Hughes et al. (2018) využili dvoufotonovou mikroskopii s kraniálními zobrazovacími okny k zobrazení hloubky až 400 μm do kůry. Podobné techniky lze použít k zobrazení povrchových myelinizovaných drah v míše v čase (Locatelli et al., 2018), ale do hlubších oblastí CNS nelze proniknout pouze dvoufotonovou mikroskopií. Jednou z alternativ je použití dvoufotonové mikroendoskopie, kdy se do tkáně zavede mikroendoskopická sonda s gradientním indexem lomu (GRIN) a zobrazí se buňky hlouběji v mozku . Zavedení endoskopu však může vést k zánětlivým reakcím, které by mohly ovlivnit myelinizaci. Alternativou by mohla být třífotonová mikroskopie s využitím metody kraniálního zobrazovacího okna, která byla dříve použita i k zobrazení hipokampu (Horton et al., 2013; Ouzounov et al., 2017). Třífotonová mikroskopie poskytuje výrazně větší poměr signálu k pozadí než dvoufotonová mikroskopie, a lze ji proto použít k zobrazení hlubších tkáňových struktur.
Zejména je důležité brát v úvahu nejen různé oblasti CNS, ale i různé neurony v těchto oblastech. Předchozí výzkum naznačuje, že existují mechanistické rozdíly v tom, jak různé podtypy neuronů regulují svou myelinizaci (Koudelka et al., 2016). Kromě toho může existovat různorodost v lokální regulaci myelinu. Je nutné si uvědomit, že různé části CNS nejsou oddělené entity, ale jsou vzájemně propojené. Integrace mezomiky, která se zaměřuje na pochopení propojení různých podtypů neuronů v různých oblastech (Zeng, 2018), bude mít zásadní význam pro naše pochopení toho, jak se liší dynamika celoživotní myelinizace v různých okruzích.
Jaký je funkční důsledek regulace myelinu podél různých okruhů? Zatím lze funkční důsledky odvodit pouze na základě korelací s chováním. V konečném důsledku je třeba spojit měření dynamiky myelinu s přímým hodnocením aktivity okruhů. To bude vyžadovat záznam neuronální aktivity během longitudinálních studií myelinizace, aby bylo možné přímo spojit pozorovanou de novo myelinizaci nebo remodelaci pochvy se změnami funkce obvodu v čase. Bude důležité měřit dynamiku myelinu a elektrofyziologickou aktivitu jednotlivých neuronů a axonů, aby bylo možné určit, jak změny různých parametrů myelinové pochvy skutečně ovlivňují vodivé vlastnosti na úrovni jednotlivých buněk, a také posoudit aktivitu na úrovni populace. Nástroje, jako jsou geneticky kódované indikátory Ca2+ nebo napětí, umožňují relativně neinvazivní záznam aktivity obvodů a lze je použít i k posouzení aktivity celých mozkových obvodů (Ahrens et al., 2012; Lovett-Barron et al., 2017).
Závěr
Myelinizace axonů představuje silný potenciální mechanismus regulace funkce obvodů v průběhu života. Výzkum prokázal, že k myelinizaci de novo v mozkové kůře (prostřednictvím produkce nových oligodendrocytů) dochází i v dospělosti a že ji lze posílit stimulací činnosti okruhu. Jakmile se myelin jednou vytvoří, je stabilní s malou obměnou oligodendrocytů a omezenou remodelací délky stávajících myelinových pochev. Tyto stabilní struktury si však mohou zachovat schopnost remodelace, pokud je myelin narušen. To má zajímavé důsledky, pokud jde o plasticitu myelinu při udržování funkce obvodů během poranění, nemoci a stáří. Jak přesně změny v myelinizaci ovlivňují funkci základního obvodu, se teprve ukáže. Nakonec je k plnému zhodnocení toho, jak dynamická myelinizace ovlivňuje celkovou funkci nervového systému v průběhu života, zapotřebí přístup na úrovni obvodu, který by integroval analýzu dynamiky myelinu s přímým měřením funkce obvodu.
Příspěvky autorů
Všichni uvedení autoři se na práci významně, přímo a intelektuálně podíleli a schválili její zveřejnění.
Financování
DAL je podporován stipendiem Wellcome Trust Senior Research Fellowship (102836/Z/13/Z). JMW je podporován grantem University of Edinburgh Ph.D. Tissue Repair Studentship Award (MRC Doctoral Training Partnership MR/K501293/1) a Wellcome Trust Four-Year Ph.D. Program in Tissue Repair (Grant 108906/Z/15/Z).
Prohlášení o střetu zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Ahrens, M. B., Li, J. M., Orger, M. B., Robson, D. N., Schier, A. F., Engert, F. a další (2012). Celomozková neuronální dynamika během motorické adaptace u zebřiček. Nature 485, 471-477. doi: 10.1038/nature11057
PubMed Abstract | CrossRef Full Text | Google Scholar
Almeida, R. G., and Lyons, D. A. (2017). O plasticitě myelinizovaných axonů a tvorbě a funkci neuronálních okruhů. J. Neurosci. 37, 10023-10034. doi: 10.1523/JNEUROSCI.3185-16.2017
CrossRef Full Text | Google Scholar
Arancibia-Cárcamo, I. L., Ford, M. C., Cossell, L., Ishida, K., Tohyama, K., and Attwell, D. (2017). Node of Ranvier length as a potential regulator of myelinated axon conduction speed. eLife 6:e23329. doi: 10.7554/eLife.23329
PubMed Abstract | CrossRef Full Text | Google Scholar
Auer, F., Vagionitis, S., and Czopka, T. (2018). Důkazy o remodelaci myelinových pochev v CNS odhalené pomocí zobrazování in vivo. Curr. Biol. 28, 549-559. doi: 10.1016/j.cub.2018.01.017
PubMed Abstract | CrossRef Full Text | Google Scholar
Barres, B. A., Hart, I. K., Coles, H. S. R., Burne, J. F., Voyvodic, J. T., Richardson, W. D., et al. (1992). Buněčná smrt a kontrola přežívání buněk v oligodendrocytární linii. Cell 70, 31-46. doi: 10.1016/0092-8674(92)90531-G
CrossRef Full Text | Google Scholar
Bechler, M. E., Byrne, L., and Ffrench-Constant, C. (2015). Délka myelinových pochev CNS je vnitřní vlastností oligodendrocytů. Curr. Biol. 25, 2411-2416. doi: 10.1016/j.cub.2015.07.056
PubMed Abstract | CrossRef Full Text | Google Scholar
Chang, A., Nishiyama, A., Peterson, J., Prineas, J., and Trapp, B. D. (2000). NG2-pozitivní oligodendrocytární progenitorové buňky v dospělém lidském mozku a v lézích roztroušené sklerózy. J. Neurosci. 20, 6404-6412. doi: 10.1523/JNEUROSCI.20-17-06404.2000
PubMed Abstract | CrossRef Full Text | Google Scholar
Cox, S. R., Ritchie, S. J., Tucker-Drob, E. M., Liewald, D. C., Hagenaars, S. P., Davies, G., et al. (2016). Stárnutí a struktura bílé hmoty mozku u 3 513 účastníků UK Biobank. Nat. Commun. 7:13629. doi: 10.1038/ncomms13629
PubMed Abstract | CrossRef Full Text | Google Scholar
Czopka, T., Ffrench-Constant, C., and Lyons, D. A. (2013). Jednotlivé oligodendrocyty mají jen několik hodin na to, aby in vivo vytvořily nové myelinové pochvy. Dev. Cell 25, 599-609. doi: 10.1016/j.devcel.2013.05.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Dawson, M. R. L., Polito, A., Levine, J. M., and Reynolds, R. (2003). Gliové progenitorové buňky exprimující NG2: hojná a rozšířená populace cyklických buněk v CNS dospělého potkana. Mol. Cell. Neurosci. 24, 476-488. doi: 10.1016/S1044-7431(03)00210-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Fard, M. K., Van der Meer, F., Sánchez, P., Cantuti-Castelvetri, L., Mandad, S., Jäkel, S., et al. (2017). Exprese BCAS1 definuje populaci časných myelinizačních oligodendrocytů v lézích roztroušené sklerózy. Sci. Transl. Med. 9:eaam7816. doi: 10.1126/scitranslmed.aam7816
PubMed Abstract | CrossRef Full Text | Google Scholar
Fields, R. D. (2015). Nový mechanismus plasticity nervového systému: myelinizace závislá na aktivitě. Nat. Rev. Neurosci. 16, 756-767. doi: 10.1038/nrn4023
PubMed Abstract | CrossRef Full Text | Google Scholar
Ford, M. C., Alexandrova, O., Cossell, L., Stange-Marten, A., Sinclair, J., Kopp-Scheinpflug, C., et al. (2015). Ladění vlastností Ranvierova uzlu a internodů v myelinizovaných axonech za účelem přizpůsobení časování akčního potenciálu. Nat. Commun. 6:8073. doi: 10.1038/ncomms9073
PubMed Abstract | CrossRef Full Text | Google Scholar
Gibson, E. M., Purger, D., Mount, C. W., Goldstein, A. K., Lin, G. L., Wood, L. S., et al. (2014). Neuronální aktivita podporuje oligodendrogenezi a adaptivní myelinizaci v mozku savců. Science 344:1252304. doi: 10.1126/science.1252304
PubMed Abstract | CrossRef Full Text | Google Scholar
Hill, R. A., Li, A. M., and Grutzendler, J. (2018). Celoživotní kortikální myelinová plasticita a degenerace související s věkem v mozku živých savců. Nat. Neurosci. 21, 683-695. doi: 10.1038/s41593-018-0120-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Hill, R. A., Patel, K. D., Goncalves, C. M., Grutzendler, J., and Nishiyama, A. (2014). Modulace tvorby oligodendrocytů během kritického časového okna po dělení NG2 buněk. Nat. Neurosci. 17, 1518-1527. doi: 10.1038/nn.3815
PubMed Abstract | CrossRef Full Text | Google Scholar
Horton, N. G., Wang, K., Kobat, D., Clark, C. G., Wise, F. W., Schaffer, C. B., et al. (2013). In vivo třífotonová mikroskopie subkortikálních struktur v intaktním myším mozku. Nat. Photonics 7, 205-209. doi: 10.1038/NPHOTON.2012.336
PubMed Abstract | CrossRef Full Text | Google Scholar
Huang, B., Wei, W., Wang, G., Gaertig, M. A., Feng, Y., Wang, W., et al. (2015). Mutantní huntingtin downreguluje expresi myelinových regulačních faktorů zprostředkovanou myelinovými geny a ovlivňuje zralé oligodendrocyty. Neuron 85, 1212-1226. doi: 10.1016/j.neuron.2015.02.026
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, E. G., Kang, S. H., Fukaya, M., and Bergles, D. E. (2013). Oligodendrocytární progenitory vyvažují růst se sebeodpuzováním, aby dosáhly homeostázy v dospělém mozku. Nat. Neurosci. 16, 668-676. doi: 10.1038/nn.3390
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., and Bergles, D. E. (2018). Remodelace myelinu prostřednictvím oligodendrogeneze závislé na zkušenosti v dospělé somatosenzorické kůře. Nat. Neurosci. 21, 696-706. doi: 10.1038/s41593-018-0121-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Jung, J. C., Mehta, A. D., Aksay, E., Stepnoski, R., and Schnitzer, M. J. (2004). Zobrazování mozku savců in vivo pomocí jedno- a dvoufotonové fluorescenční mikroendoskopie. J. Neurophysiol. 92, 3121-3133. doi: 10.1152/jn.00234.2004
PubMed Abstract | CrossRef Full Text | Google Scholar
Kang, S. H., Li, Y., Fukaya, M., Lorenzini, I., Cleveland, D. W., Ostrow, L. W., et al. (2013). Degenerace a porucha regenerace oligodendrocytů šedé hmoty u amyotrofické laterální sklerózy. Nat. Neurosci. 16, 571-579. doi: 10.1038/nn.3357
PubMed Abstract | CrossRef Full Text | Google Scholar
Koudelka, S., Voas, M. G., Almeida, R. G., Baraban, M., Soetaert, J., Meyer, M. P., et al. (2016). Jednotlivé podtypy neuronů vykazují rozmanitost v myelinizaci CNS zprostředkované uvolňováním synaptických vezikul. Curr. Biol. 26, 1447-1455. doi: 10.1016/j.cub.2016.03.070
PubMed Abstract | CrossRef Full Text | Google Scholar
Kougioumtzidou, E., Shimizu, T., Hamilton, N. B., Tohyama, K., Sprengel, R., Monyer, H., et al. (2017). Signalizace prostřednictvím receptorů AMPA na prekurzorech oligodendrocytů podporuje myelinizaci tím, že zvyšuje přežívání oligodendrocytů. eLife 6:e28080. doi: 10.7554/eLife.28080
PubMed Abstract | CrossRef Full Text | Google Scholar
Kwon, J., Kim, M., Park, H., Kang, B.-M., Jo, Y., Kim, J.-H., et al. (2017). Optická metrologie bez označení v nanorozměrech na myelinizovaných axonech in vivo. Nat. Commun. 8:1832. doi: 10.1038/s41467-017-01979-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Levene, M. J., Dombeck, D. A., Kasischke, K. A., Molloy, R. P., and Webb, W. W. (2004). Multifotonová mikroskopie hluboké mozkové tkáně in vivo. J. Neurophysiol. 91, 1908-1912. doi: 10.1152/jn.01007.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Lim, H., Sharoukhov, D., Kassim, I., Zhang, Y., Salzer, J. L., and Melendez-Vasquez, C. V. (2014). Zobrazování myelinizace Schwannových buněk bez použití značek pomocí mikroskopie třetí harmonické generace. Proc. Natl. Acad. Sci. U.S.A. 111, 18025-18030. doi: 10.1073/pnas.1417820111
PubMed Abstract | CrossRef Full Text | Google Scholar
Locatelli, G., Theodorou, D., Kendirli, A., Jordão, M. J. C., Staszewski, O., Phulphagar, K., et al. (2018). Mononukleární fagocyty lokálně specifikují a přizpůsobují svůj fenotyp v modelu roztroušené sklerózy. Nat. Neurosci. 21, 1196-1208. doi: 10.1038/s41593-018-0212-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovett-Barron, M., Andalman, A. S., Allen, W. E., Vesuna, S., Kauvar, I., Burns, V. M., et al. (2017). Předpotopní obvody pro koordinovanou modulaci stavu mozku. Cell 171, 1411-1423. doi: 10.1016/j.cell.2017.10.021
PubMed Abstract | CrossRef Full Text | Google Scholar
Mabbott, D. J., Noseworthy, M., Bouffet, E., Laughlin, S., and Rockel, C. (2006). Růst bílé hmoty jako mechanismus kognitivního vývoje u dětí. Neuroimage 33, 936-946. doi: 10.1016/j.neuroimage.2006.07.024
PubMed Abstract | CrossRef Full Text | Google Scholar
McKenzie, I. A., Ohayon, D., Li, H., Paes de Faria, J., Emery, B., Tohyama, K. a další (2014). Učení motorických dovedností vyžaduje aktivní centrální myelinizaci. Science 346, 318-322. doi: 10.1126/science.1254960
PubMed Abstract | CrossRef Full Text | Google Scholar
Mitew, S., Gobius, I., Fenlon, L. R., McDougall, S. J., Hawkes, D., Xing, Y. L., et al. (2018). Farmakogenetická stimulace neuronální aktivity zvyšuje myelinizaci způsobem specifickým pro axon. Nat. Commun. 9:306. doi: 10.1038/s41467-017-02719-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Mount, C. W., and Monje, M. (2017). Wrapped to adapt: experience-dependent myelination (Zabalení k adaptaci: myelinizace závislá na zkušenostech). Neuron 95, 743-756. doi: 10.1016/j.neuron.2017.07.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Ouzounov, D. G., Wang, T., Wang, M., Feng, D. D., Horton, N. G., Cruz-Hernández, J. C., et al. (2017). In vivo třífotonové zobrazování aktivity neuronů značených GCaMP6 hluboko v intaktním myším mozku. Nat. Methods 14, 388-390. doi: 10.1038/nmeth.4183
PubMed Abstract | CrossRef Full Text | Google Scholar
Ritchie, S. J., Bastin, M. E., Tucker-Drob, E. M., Maniega, S. M., Engelhardt, L. E., Cox, S. R., et al. (2015). Propojené změny mikrostruktury bílé hmoty mozku a fluidní inteligence v pozdějším věku. J. Neurosci. 35, 8672-8682. doi: 10.1523/JNEUROSCI.0862-15.2015
PubMed Abstract | CrossRef Full Text | Google Scholar
Rivers, L. E., Young, K. M., Rizzi, M., Jamen, F., Psachoulia, K., Wade, A., et al. (2008). Glie PDGFRA/NG2 generují myelinizační oligodendrocyty a piriformní projekční neurony u dospělých myší. Nat. Neurosci. 11, 1392-1401. doi: 10.1038/nn.2220
PubMed Abstract | CrossRef Full Text | Google Scholar
Sampaio-Baptista, C., Khrapitchev, A. A., Foxley, S., Schlagheck, T., Scholz, J., Jbabdi, S., et al. (2013). Učení motorických dovedností vyvolává změny v mikrostruktuře bílé hmoty a myelinizaci. J. Neurosci. 33, 19499-19503. doi: 10.1523/jneurosci.3048-13.2013
PubMed Abstract | CrossRef Full Text | Google Scholar
Scantlebury, N., Cunningham, T., Dockstader, C., Laughlin, S., Gaetz, W., Rockel, C., et al. (2014). Vztah mezi zráním bílé hmoty a reakční dobou v dětství. J. Int. Neurophsychol. Soc. 20, 99-112. doi: 10.1017/S1355617713001148
PubMed Abstract | CrossRef Full Text | Google Scholar
Schain, A. J., Hill, R. A., and Grutzendler, J. (2014). Zobrazování myelinizovaných axonů in vivo bez označení ve zdraví a nemoci pomocí spektrální konfokální reflektanční mikroskopie. Nat. Med. 20, 443-449. doi: 10.1038/nm.3495
PubMed Abstract | CrossRef Full Text | Google Scholar
Scholz, J., Klein, M. C., Behrens, T. E. J., and Johansen-berg, H. (2009). Trénink vyvolává změny v architektuře bílé hmoty. Nat. Neurosci. 12, 1370-1371. doi: 10.1038/nn.2412
PubMed Abstract | CrossRef Full Text | Google Scholar
Snaidero, N., Möbius, W., Czopka, T., Hekking, L. H. P., Mathisen, C., Verkleij, D., et al. (2014). Obalení axonů CNS myelinovou membránou pomocí polarizovaného růstu závislého na PI(3,4,5)P3 na vnitřním jazyku. Cell 156, 277-290. doi: 10.1016/j.cell.2013.11.044
PubMed Abstract | CrossRef Full Text | Google Scholar
Takahashi, N., Sakurai, T., Davis, K. L., and Buxbaum, J. D. (2011). Spojení dysfunkce oligodendrocytů a myelinu s abnormalitami neurocircuitry u schizofrenie. Prog. Neurobiol. 93, 13-24. doi: 10.1016/j.pneurobio.2010.09.004
PubMed Abstract | CrossRef Full Text | Google Scholar
Tomassy, G. S., Berger, D. R., Chen, H.-H., Kasthuri, N., Hayworth, K. J., Vercelli, A., et al. (2014). Odlišné profily distribuce myelinu podél jednotlivých axonů pyramidových neuronů v neokortexu. Science 344, 319-324. doi: 10.1126/science.1249766
PubMed Abstract | CrossRef Full Text | Google Scholar
Tripathi, R. B., Jackiewicz, M., Mckenzie, I. A., Kougioumtzidou, E., Grist, M., and Richardson, W. D. (2017). Pozoruhodná stabilita myelinizačních oligodendrocytů u myší. Cell Rep. 21, 316-323. doi: 10.1016/j.celrep.2017.09.050
PubMed Abstract | CrossRef Full Text | Google Scholar
Viganò, F., Möbius, W., Götz, M., and Dimou, L. (2013). Transplantace odhaluje regionální rozdíly v diferenciaci oligodendrocytů v dospělém mozku. Nat. Neurosci. 16, 1370-1372. doi: 10.1038/nn.3503
PubMed Abstract | CrossRef Full Text | Google Scholar
Watkins, T. A., Emery, B., Mulinyawe, S., and Barres, B. A. (2008). Odlišné fáze myelinizace regulované gama-sekretázou a astrocyty v rychle myelinizujícím kokulturním systému CNS. Neuron 60, 555-569. doi: 10.1016/j.neuron.2008.09.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Waxman, S. G. (1980). Determinanty rychlosti vedení v myelinizovaných nervových vláknech. Muscle Nerve 3, 141-150. doi: 10.1002/mus.880030207
PubMed Abstract | CrossRef Text | Google Scholar
Xiao, L., Ohayon, D., Mckenzie, I. A., Sinclair-Wilson, A., Wright, J. L., Fudge, A. D., et al. (2016). Rychlá tvorba nových oligodendrocytů je nutná v nejranějších fázích učení motorických dovedností. Nat. Neurosci. 19, 1210-1217. doi: 10.1038/nn.4351
PubMed Abstract | CrossRef Full Text | Google Scholar
Yeung, M. S. Y., Zdunek, S., Bergmann, O., Bernard, S., Salehpour, M., Alkass, K., et al. (2014). Dynamika tvorby oligodendrocytů a myelinizace v lidském mozku. Cell 159, 766-774. doi: 10.1016/j.cell.2014.10.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Young, K. M., Psachoulia, K., Tripathi, R. B., Dunn, S.-J., Cossell, L., Attwell, D., et al. (2013). Oligodendrocytární dynamika ve zdravém CNS dospělých: důkaz remodelace myelinu. Neuron 77, 873-885. doi: 10.1016/j.neuron.2013.01.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Zeng, H. (2018). Mesoscale connectomics. Curr. Opin. Neurobiol. 50, 154-162. doi: 10.1016/j.conb.2018.03.003
PubMed Abstract | CrossRef Full Text | Google Scholar
.