Excitation-contraction coupling
Excitation-contraction coupling je spojení (transdukce) mezi akčním potenciálem generovaným v sarkolemě a začátkem svalové kontrakce. Spouštěčem uvolňování vápníku ze sarkoplazmatického retikula do sarkoplazmy je nervový signál. Každé vlákno kosterního svalu je ovládáno motorickým neuronem, který vede signály z mozku nebo míchy do svalu. Oblast sarkolemmy na svalovém vlákně, která interaguje s neuronem, se nazývá koncová destička motoru. Konec axonu neuronu se nazývá synaptický terminál a ve skutečnosti se motorické koncové destičky nedotýká. Synaptický terminál od motorické koncové destičky odděluje malý prostor zvaný synaptická štěrbina. Elektrické signály se šíří podél axonu neuronu, který se větví ve svalu a připojuje se k jednotlivým svalovým vláknům v nervosvalovém spojení.
Schopnost buněk elektricky komunikovat vyžaduje, aby buňky vynaložily energii na vytvoření elektrického gradientu přes své buněčné membrány. Tento gradient náboje je přenášen ionty, které jsou v membráně různě rozmístěny. Každý iont působí elektrickým vlivem a vlivem koncentrace. Stejně jako se mléko nakonec smísí s kávou bez nutnosti míchání, i ionty se také rovnoměrně rozdělí, pokud je jim to umožněno. V tomto případě jim není dovoleno vrátit se do rovnoměrně promíchaného stavu.
Natrium-draslíková ATPáza využívá buněčnou energii k přesunu iontů K+ dovnitř buňky a iontů Na+ ven. To samo o sobě akumuluje malý elektrický náboj, ale velký koncentrační gradient. V buňce je hodně K+ a mimo buňku hodně Na+. Draslík může buňku opustit přes K+ kanály, které jsou v 90 % případů otevřené, a také to dělá. Na+ kanály jsou však otevřeny jen zřídka, takže Na+ zůstává mimo buňku. Když K+ opouští buňku a řídí se koncentračním gradientem, zanechává za sebou záporný náboj. V klidovém stavu tedy existuje velký koncentrační gradient pro vstup Na+ do buňky a v buňce se hromadí záporné náboje. To je klidový membránový potenciál. Potenciál v tomto kontextu znamená oddělení elektrického náboje, který je schopen konat práci. Měří se ve voltech, stejně jako u baterie. Transmembránový potenciál je však podstatně menší (0,07 V); proto se malá hodnota vyjadřuje jako milivolty (mV) nebo 70 mV. Protože vnitřek buňky je ve srovnání s vnějškem záporný, znaménko minus znamená přebytek záporných nábojů uvnitř buňky, tedy -70 mV.
Pokud nějaká událost změní propustnost membrány pro ionty Na+, proniknou do buňky. Tím se změní napětí. Jedná se o elektrickou událost, nazývanou akční potenciál, kterou lze využít jako buněčný signál. Ke komunikaci mezi nervy a svaly dochází prostřednictvím neurotransmiterů. Akční potenciály neuronů způsobují uvolňování neurotransmiterů ze synaptického terminálu do synaptické štěrbiny, kde pak mohou difundovat přes synaptickou štěrbinu a vázat se na molekulu receptoru na koncové destičce motoru. Motorická koncová destička má junkční záhyby – záhyby v sarkolemě, které vytvářejí velký povrch pro vazbu neurotransmiteru na receptory. Receptory jsou vlastně sodíkové kanály, které se po přijetí signálu neurotransmiteru otevírají a umožňují průchod Na+ do buňky.
Acetylcholin (ACh) je neurotransmiter uvolňovaný motorickými neurony, který se váže na receptory v koncové destičce motoru. K uvolnění neurotransmiteru dochází při průchodu akčního potenciálu axonem motorického neuronu, což vede ke změně propustnosti membrány synaptického zakončení a přílivu vápníku. Ionty Ca2+ umožňují synaptickým vezikulám přesun a spojení s presynaptickou membránou (na neuronu) a uvolnění neurotransmiteru z vezikul do synaptické štěrbiny. Po uvolnění synaptickým terminálem ACh difunduje přes synaptickou štěrbinu k motorické koncové destičce, kde se váže s ACh receptory. Při vazbě neurotransmiteru se tyto iontové kanály otevřou a ionty Na+ projdou membránou do svalové buňky. Tím se sníží rozdíl napětí mezi vnitřní a vnější stranou buňky, což se nazývá depolarizace. Protože se ACh váže na koncové destičce motoru, nazývá se tato depolarizace potenciál koncové destičky. Depolarizace se pak šíří podél sarkolemy a vytváří akční potenciál, protože sodíkové kanály sousedící s místem počáteční depolarizace detekují změnu napětí a otevřou se. Akční potenciál se pohybuje napříč celou buňkou a vytváří vlnu depolarizace.
ACh je rozkládán enzymem acetylcholinesterázou (AChE) na acetyl a cholin. AChE se nachází v synaptické štěrbině a rozkládá ACh tak, aby nezůstal vázán na ACh receptory, což by způsobilo nežádoucí prodlouženou svalovou kontrakci (obr. 6.9).
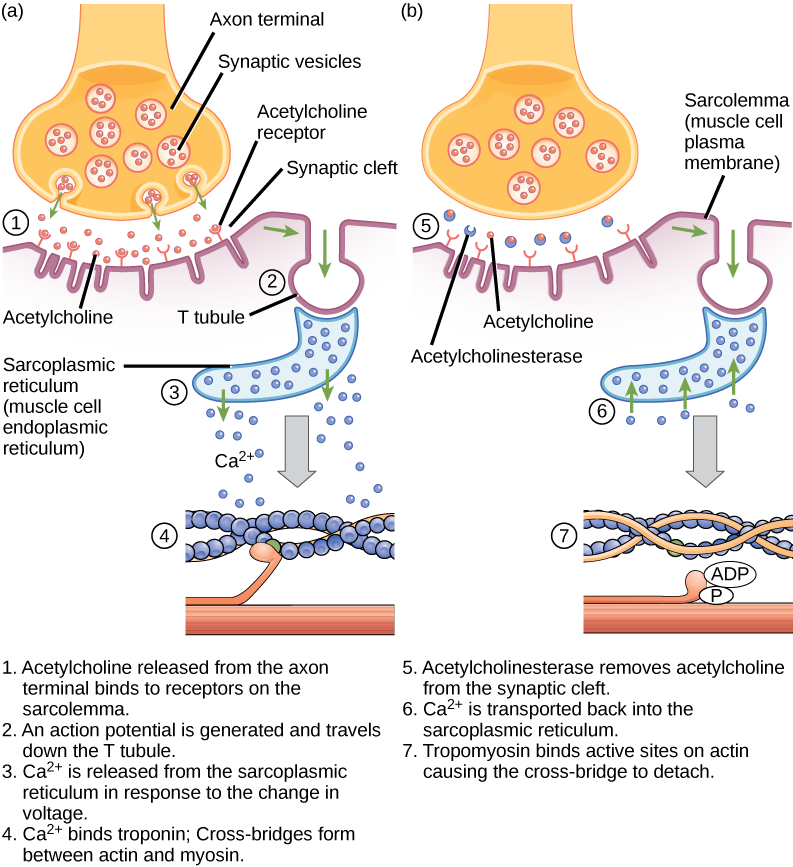
Obrázek 6.9. Toto schéma znázorňuje vazbu mezi vzruchem a kontrakcí při kontrakci kosterního svalu. Sarkoplazmatické retikulum je specializované endoplazmatické retikulum, které se nachází ve svalových buňkách.
Cvičení \(\PageIndex{1}\)
Smrtící nervový plyn sarin nevratně inhibuje acetylcholinesterázu. Jaký vliv by měl Sarin na svalovou kontrakci?
Po depolarizaci se membrána vrátí do klidového stavu. Tomu se říká repolarizace, během níž se uzavírají napěťově řízené sodíkové kanály. Draslíkové kanály pokračují při 90% vodivosti. Protože sodíko-draslíková ATPáza v plazmatické membráně vždy transportuje ionty, obnoví se klidový stav (záporně nabitá vnitřní strana vzhledem k vnější). Období bezprostředně následující po přenosu impulzu v nervu nebo svalu, kdy neuron nebo svalová buňka znovu získávají schopnost přenášet další impuls, se nazývá refrakterní perioda. Během refrakterní periody nemůže membrána generovat další akční potenciál. Refrakterní perioda umožňuje iontovým kanálům citlivým na napětí vrátit se do své klidové konfigurace. Sodíko-draslíková ATPáza neustále přesouvá Na+ zpět z buňky a K+ zpět do buňky a K+ uniká ven a zanechává za sebou záporný náboj. Membrána se velmi rychle repolarizuje, takže může být opět depolarizována.