Excitation-kontraktionskoppling
Excitation-kontraktionskoppling är länken (transduktionen) mellan den aktionspotential som genereras i sarkolemma och starten av en muskelkontraktion. Den utlösande faktorn för kalciumfrisättning från det sarkoplasmatiska retikulumet till sarkoplasman är en neuralsignal. Varje skelettmuskelfiber styrs av ett motorneuron, som leder signaler från hjärnan eller ryggmärgen till muskeln. Det område av sarkolemman på muskelfibrerna som interagerar med neuronet kallas den motoriska ändplattan. Änden av neuronens axon kallas den synaptiska terminalen, och den kommer faktiskt inte i kontakt med den motoriska ändplattan. Ett litet utrymme som kallas den synaptiska klyftan skiljer den synaptiska terminalen från den motoriska ändplattan. Elektriska signaler färdas längs neuronens axon, som förgrenar sig genom muskeln och ansluter till enskilda muskelfibrer vid en neuromuskulär korsning.
Förmågan hos celler att kommunicera elektriskt kräver att cellerna förbrukar energi för att skapa en elektrisk gradient över sina cellmembran. Denna laddningsgradient bärs upp av joner som är differentiellt fördelade över membranet. Varje jon utövar ett elektriskt inflytande och ett koncentrationsinflytande. Precis som mjölk så småningom blandas med kaffe utan att man behöver röra om, fördelar jonerna sig också jämnt, om de tillåts göra det. I det här fallet tillåts de inte att återgå till ett jämnt blandat tillstånd.
Natrium-kalium ATPaset använder cellenergi för att flytta K+-joner inuti cellen och Na+-joner utanför. Enbart detta ackumulerar en liten elektrisk laddning, men en stor koncentrationsgradient. Det finns mycket K+ i cellen och mycket Na+ utanför cellen. Kalium kan lämna cellen genom K+-kanaler som är öppna 90 procent av tiden, och det gör det. Na+-kanalerna är dock sällan öppna, så Na+ stannar utanför cellen. När K+ lämnar cellen och följer sin koncentrationsgradient lämnar det i själva verket en negativ laddning kvar. I vila finns det alltså en stor koncentrationsgradient för Na+ att komma in i cellen, och det finns en ackumulering av negativa laddningar som lämnas kvar i cellen. Detta är den vilande membranpotentialen. Med potential menas i detta sammanhang en separation av elektrisk laddning som kan utföra arbete. Den mäts i volt, precis som i ett batteri. Transmembranpotentialen är dock betydligt mindre (0,07 V); därför uttrycks det lilla värdet som millivolt (mV) eller 70 mV. Eftersom insidan av en cell är negativ jämfört med utsidan innebär ett minustecken överskottet av negativa laddningar inuti cellen, -70 mV.
Om en händelse förändrar membranets genomsläpplighet för Na+-joner kommer de in i cellen. Det kommer att förändra spänningen. Detta är en elektrisk händelse, kallad aktionspotential, som kan användas som en cellsignal. Kommunikation sker mellan nerver och muskler genom neurotransmittorer. Neuronernas aktionspotential orsakar frisättning av neurotransmittorer från den synaptiska terminalen till den synaptiska klyftan, där de sedan kan diffundera genom den synaptiska klyftan och binda till en receptormolekyl på den motoriska ändplattan. Den motoriska ändplattan har junctional folds – veck i sarkolemman som skapar en stor yta där neurotransmittorn kan binda till receptorerna. Receptorerna är i själva verket natriumkanaler som öppnas för att låta Na+ passera in i cellen när de tar emot signalsubstansen från neurotransmittorn.
Acetylkolin (ACh) är en neurotransmittor som frisätts av motoriska neuroner och som binder till receptorer i den motoriska ändplattan. Frisättning av neurotransmittor sker när en aktionspotential färdas ner i motorneuronets axon, vilket resulterar i förändrad permeabilitet i det synaptiska terminalmembranet och ett inflöde av kalcium. Ca2+-jonerna gör det möjligt för synaptiska vesiklar att förflytta sig till och binda sig till det presynaptiska membranet (på neuronen) och frigöra neurotransmittor från vesiklarna till den synaptiska klyftan. När ACh frisätts av den synaptiska terminalen diffunderar det genom den synaptiska klyftan till den motoriska ändplattan, där det binder till ACh-receptorer. När en neurotransmittor binder sig öppnas dessa jonkanaler och Na+-joner passerar membranet in i muskelcellen. Detta minskar spänningsskillnaden mellan cellens insida och utsida, vilket kallas depolarisering. Eftersom ACh binder vid den motoriska ändplattan kallas denna depolarisering för en ändplattpotential. Depolariseringen sprider sig sedan längs sarkolemman och skapar en aktionspotential när natriumkanaler intill den ursprungliga depolariseringsplatsen upptäcker spänningsförändringen och öppnas. Aktionspotentialen rör sig över hela cellen och skapar en våg av depolarisering.
ACh bryts ner av enzymet acetylkolinesteras (AChE) till acetyl och kolin. AChE finns i den synaptiska klyftan och bryter ner ACh så att det inte förblir bundet till ACh-receptorer, vilket skulle orsaka oönskade förlängda muskelkontraktioner (figur 6.9).
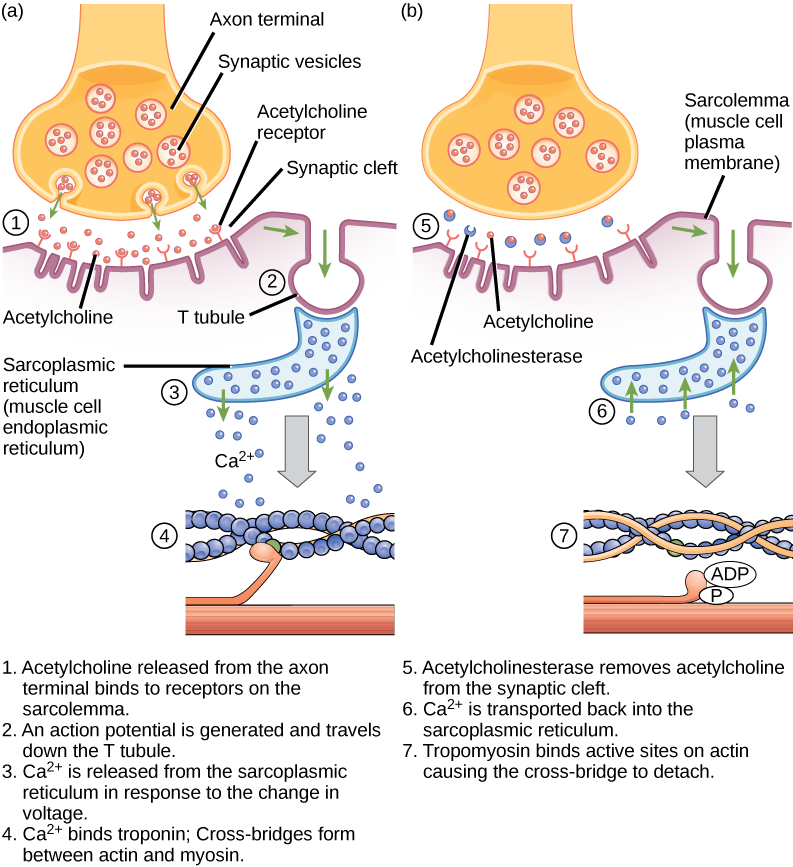
Figur 6.9. Detta diagram visar excitation-kontraktionskopplingen i en skelettmuskelkontraktion. Det sarkoplasmatiska retikulumet är ett specialiserat endoplasmatiskt retikulum som finns i muskelceller.
Träning \(\PageIndex{1}\)
Den dödliga nervgasen Sarin hämmar acetylkolinesteras irreversibelt. Vilken effekt skulle Sarin ha på muskelkontraktion?
Efter depolarisering återgår membranet till sitt viloläge. Detta kallas repolarisering, under vilken spänningsstyrda natriumkanaler stängs. Kaliumkanalerna fortsätter med 90 % konduktivitet. Eftersom plasmamembranets natrium-kalium ATPas alltid transporterar joner återställs vilotillståndet (negativt laddad insida i förhållande till utsidan). Den period som följer omedelbart efter överföringen av en impuls i en nerv eller muskel, under vilken en neuron eller muskelcell återfår sin förmåga att överföra en annan impuls, kallas refraktärperioden. Under den refraktära perioden kan membranet inte generera en ny aktionspotential. Refraktärperioden gör det möjligt för de spänningskänsliga jonkanalerna att återgå till sina vilokonfigurationer. Natrium-kalium ATPaset flyttar kontinuerligt Na+ tillbaka ut ur cellen och K+ tillbaka in i cellen, och K+ läcker ut och lämnar negativ laddning kvar. Mycket snabbt repolariseras membranet, så att det återigen kan depolariseras.