Introducere
Creierul uman suferă o maturizare extinsă pe parcursul vieții pentru a facilita dezvoltarea cognitivă. Mielinizarea axonilor din întregul sistem nervos este unul dintre aceste procese cruciale de maturizare. În sistemul nervos central (SNC), celulele gliale numite oligodendrocite extind numeroase procese în mediul lor înconjurător, care înfășoară concentric membrana în jurul axonilor pentru a forma teci de mielină. Învelișurile de mielină permit conducerea saltatorie rapidă a potențialelor de acțiune, prin localizarea canalelor Na+ activate de tensiune în spațiile scurte dintre învelișurile adiacente (cunoscute sub numele de nodurile Ranvier) și prin faptul că acționează ca izolatori electrici. Axonii care sunt complet mielinizați pe toată lungimea lor conduc impulsurile de multe ori mai repede decât axonii nemielinizați cu aceeași dimensiune a secțiunii transversale (Waxman, 1980). Prin urmare, circuitele neuronale mielinizate conduc informațiile mult mai rapid decât circuitele nemielinizate. Oamenii se nasc cu un SNC practic nemielinizat, iar populația de oligodendrocite se extinde dramatic după naștere, cu mielinizare pe scară largă în primii câțiva ani ai copilăriei. Mielinizarea continuă în adolescență și la vârsta adultă într-o manieră spațio-temporală caracteristică, în corelație cu apariția și menținerea unei funcții adecvate a circuitelor. De exemplu, maturizarea materiei albe (zonele bogate în mielină din SNC) este concomitentă cu dezvoltarea proceselor cognitive din copilărie, cum ar fi viteza de procesare a informațiilor (Mabbott et al., 2006; Scantlebury et al., 2014). În plus, patologia/anomalii ale mielinei sunt observate nu numai în boala demielinizantă scleroza multiplă, ci și în mai multe boli neurodegenerative (Kang et al., 2013; Huang et al., 2015) și tulburări de neurodezvoltare (Takahashi et al., 2011). Cu toate acestea, mielinizarea axonilor individuali nu este un fenomen „totul sau nimic”. Axonii din SNC prezintă variații extinse în ceea ce privește numărul de teci de mielină, lungimea tecii, grosimea tecii și distribuția de-a lungul lungimii lor. Există multe modele diferite de mielinizare; de exemplu, axonii cu regiuni slab mielinizate au fost descrise în cortexul de șoarece tânăr și adult (Tomassy et al., 2014; Hill et al., 2018; Hughes et al., 2018). Modificarea oricăruia dintre acești parametri ai tecii are efecte previzibile asupra vitezei de conducere a axonului subiacent – prin urmare, stabilirea unui model specific de mielinizare de-a lungul unui axon poate fi deosebit de importantă pentru reglarea fină a funcției circuitului. De exemplu, axonii din trunchiul cerebral auditiv al gerbililor prezintă învelișuri de mielină din ce în ce mai scurte de-a lungul regiunilor distale pentru a asigura sincronizarea precisă a sosirii semnalului pentru a facilita localizarea sunetului (Ford et al., 2015). Modificări subtile ale modelului general de mielinizare de-a lungul unui axon (fie prin adăugarea de mielină nouă, fie prin remodelarea celei existente) ar putea schimba profund sincronizarea impulsurilor neuronale în circuite. Dacă mielina este adaptabilă, atunci modificarea unor astfel de modele de mielinizare ar putea reprezenta un mecanism puternic de reglare a funcției circuitelor pe tot parcursul vieții.
Povezi recente sugerează că mielina poate fi adaptabilă ca răspuns la activitatea circuitului. Imagistica tensorială de difuzie la nivelul întregului creier poate fi utilizată pentru a măsura modificările ample ale materiei albe bogate în mielină de-a lungul timpului – astfel de experimente la oameni și rozătoare au arătat că învățarea unei sarcini noi se corelează cu modificări ale materiei albe în regiuni relevante ale creierului (Scholz et al., 2009; Sampaio-Baptista et al., 2013). Analizele la nivel celular în modele animale demonstrează că producția de noi oligodendrocite mielinizante este necesară pentru o învățare motorie eficientă (McKenzie et al., 2014; Xiao et al., 2016). În prezent, se presupune că activitatea circuitelor neuronale poate declanșa modificări ale mielinei; un corpus extins de cercetări a demonstrat că activitatea neuronală poate influența proliferarea celulelor precursoare ale oligodendrocitelor (OPC), diferențierea oligodendrocitelor, precum și formarea și creșterea tecilor de mielină. Aceste cercetări, inclusiv dovezile privind semnalele moleculare implicate, au fost analizate pe larg în altă parte (Fields, 2015; Almeida și Lyons, 2017; Mount și Monje, 2017). Activitatea neuronală ar putea determina modificări ale mielinei care, la rândul său, ar putea modifica vitezele de conducere pentru a regla cu precizie temporizările care stau la baza funcționării circuitelor.
Cu toate acestea, încă nu știm dacă sau cum este reglementată în mod dinamic mielinizarea circuitelor de-a lungul vieții. Lucrările la rozătoare indică faptul că noi oligodendrocite sunt generate în întregul SNC chiar și la vârsta adultă (Young et al., 2013), iar OPC-urile rezidă în creierul uman adult (Chang et al., 2000). Analiza datarea cu carbon a țesuturilor umane a identificat oligodendrocite născute la vârsta adultă în cortex, deși aceleași analize au indicat că majoritatea oligodendrocitelor din corpul calos își au originea în copilăria timpurie (Yeung et al., 2014). Cu toate acestea, studiile de neuroimagistică la om care corelează alterările structurale ale materiei albe cu învățarea sarcinilor sugerează că noua mielină se poate forma pe tot parcursul vieții. O astfel de mielinizare prelungită ar necesita, în principiu, o producție de oligodendrocite pe tot parcursul vieții, având în vedere că oligodendrocitele individuale de mielinizare au o fereastră de timp restrânsă de doar câteva ore pentru a iniția formarea de noi teci (Watkins et al., 2008; Czopka et al., 2013), iar numărul de teci pe oligodendrocit pare stabil în timp (Tripathi et al., 2017). O avertizare observată de Mount și Monje (2017) este că data „nașterii” în experimentul de datare cu carbon (care identifică momentul replicării ADN-ului în timpul diviziunii celulare), o reflectă pe cea a OPC, nu neapărat oligodendrocitele diferențiate. Acest lucru este important având în vedere dovezile conform cărora OPC-urile se pot diferenția direct în oligodendrocite fără diviziune celulară, cel puțin la rozătoare (Hughes et al., 2013). OPC-urile din corpul calos ar putea să se diferențieze direct în oligodendrocite la mulți ani după diviziunea lor celulară terminală; astfel, momentul diferențierii acestor noi oligodendrocite nu poate fi determinat prin datarea cu carbon și, prin urmare, este posibil ca Yeung et al. (2014) să fi subestimat rata de producere a oligodendrocitelor în creierul uman adult. Mai avem încă multe de învățat despre contribuțiile relative ale generării oligodendrocitelor și remodelării mielinei la dezvoltarea SNC de-a lungul vieții.
Pentru a înțelege pe deplin dinamica precisă a oligodendrogenezei, a formării de mielină și a remodelării mielinei de-a lungul diferitelor etape ale vieții, imagistica longitudinală de înaltă rezoluție reprezintă abordarea standard de aur. Aici oferim o prezentare generală a studiilor recente de imagistică in vivo care încep să clarifice dinamica mielinizării, ceea ce ne va permite, de asemenea, să începem să înțelegem modul în care o astfel de dinamică ar putea avea un impact asupra funcției circuitelor neuronale.
De novo Myelination
Pentru a începe să abordăm definitiv modul în care sunt generate oligodendrocitele și modul în care mielina este fabricată și remodelată dinamic in vivo, două studii recente au utilizat imagistica repetată cu doi fotoni a cortexului somatosenzorial de șoarece pe perioade extinse de timp. Hughes et al. (2018) au imaginat cortexul șoarecilor reporteri transgenici cu celule din linia oligodendroglială marcate fluorescent de la începutul vârstei adulte, până la vârsta mijlocie și bătrânețe (aproximativ P720). Aceștia au constatat că populația de oligodendrocite continuă să se extindă și că densitatea oligodendrocitelor corticale aproape se dublează între stadiile de adult tânăr și de vârstă mijlocie (Figura 1A). Acest lucru a fost însoțit de o creștere de peste două ori a numărului de teci de mielină corticale. Dar cum crește numărul oligodendrocitelor? În dezvoltarea postnatală timpurie sunt produse multe oligodendrocite, dar numai un subset supraviețuiește și continuă să mielinizeze axonii (Barres et al., 1992). Acest lucru pare să fie similar la vârsta adultă – urmărind OPC-uri corticale individuale în cortexul adult timp de până la 50 de zile, Hughes et al. (2018) au arătat că majoritatea oligodendrocitelor nou diferențiate suferă moarte celulară, doar 22% supraviețuind și angajându-se în mielinizare (Figura 1B). Rămâne necunoscută proporția de oligodendrocite nou diferențiate care sunt generate în urma diviziunii OPC față de diferențierea directă. Cu toate acestea, odată ce oligodendrocitele se angajează în mielinizare, ele rămân stabile, neexistând nicio dovadă că oligodendrocitele mielinizante suferă de moarte celulară în timpul unei perioade de imagistică de 50 de zile.
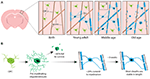
FIGURA 1. Dinamica oligodendrocitelor și a mielinei în cortexul mamiferelor de-a lungul vieții. (A) Celulele precursoare ale oligodendrocitelor (OPC) generează continuu noi oligodendrocite mielinizante (OL) în cortexul somatosenzorial de la naștere până la vârsta mijlocie. Populația de OL scade apoi la bătrânețe, însoțită de o reducere a acoperirii cu mielină. (B) Urmărirea traseului liniei de linie a unor OPC-uri individuale arată că, deși OL premelinizante sunt produse în mod continuu la vârsta adultă, doar aproximativ 20 % dintre acestea supraviețuiesc până la mielinizare. Majoritatea învelișurilor de mielină, odată formate, au o lungime stabilă pe o perioadă prelungită de timp, ceea ce indică faptul că, în mod normal, există o remodelare foarte mică a mielinei existente. Rezumat al datelor din Hill et al. (2018) și Hughes et al. (2018).
În mod similar, Hill et al. (2018) au folosit reporteri transgenici ai oligodendrocitelor și tehnica de microscopie confocală de reflectanță spectrală fără etichetă (SCoRe) pentru a vizualiza mielina de-a lungul axonilor (Schain et al., 2014) în cortexul somatosenzorial al șoarecilor tineri, adulți tineri, de vârstă mijlocie și de vârstă înaintată (P950). Aceștia au constatat, de asemenea, că numărul oligodendrocitelor continuă să se extindă la vârsta adultă până la P650 și că oligodendrocitele sunt stabile la vârsta mijlocie până la 80 de zile de imagistică. Aceștia au constatat că mielinizarea cortexului atinge, de asemenea, un vârf la vârsta mijlocie, la P650, și că densitatea oligodendrocitelor scade semnificativ de la vârf (la P650) până la vârsta foarte înaintată (P950) (Figura 1A). Acest lucru s-a reflectat într-o reducere a acoperirii cu mielină a axonilor corticali de strat I între P650 și P950. Supraviețuirea oligodendrocitelor pe termen lung poate varia între diferite părți ale SNC. Tripathi et al. (2017) au marcat oligodendrocitele mielinizante la P60 la șoareci și apoi au numărat câte celule marcate au supraviețuit până la P605 în mai multe regiuni ale SNC. Ei au constatat că în măduva spinării și cortexul motor, 60-70% dintre celulele marcate la P60 au supraviețuit, în timp ce în corpul calos, peste 90% dintre celulele marcate la P60 au supraviețuit. Reducerea numărului de oligodendrocite și a mielinizării în anumite regiuni ale SNC odată cu înaintarea în vârstă ridică întrebări intrigante cu privire la rolul pierderii de mielină în declinul cognitiv asociat cu vârsta. Analiza RMN arată că microstructura materiei albe se corelează cu inteligența fluidă (Ritchie et al., 2015), dar și că această microstructură a materiei albe se deteriorează odată cu înaintarea în vârstă (Cox et al., 2016). Pierderea ulterioară de mielină asociată vârstei ar putea duce la reducerea funcției cognitive din cauza dereglementării circuitelor mielinizate.
Ar putea ca generarea de noi oligodendrocite (și, ulterior, de nouă mielină) în cortexul adult să fie receptivă la activitatea circuitului? Cercetările anterioare au arătat că reducerea aportului senzorial prin îndepărtarea mustăților la șoareci duce la o oligodendrogeneză redusă în cortexul somatosenzorial (Hill et al., 2014). Pentru a investiga acest lucru în continuare, Hughes și colab. (2018) au oferit șoarecilor adulți (P365) stimulare senzorială timp de 3 săptămâni prin agățarea de mărgele în cuștile animalelor pentru a le stimula în mod repetat mustățile și, prin urmare, cortexul somatosenzorial. Prin imagistica cortexului somatosenzorial înainte și după cele 3 săptămâni, aceștia au demonstrat că stimularea senzorială crește numărul de oligodendrocite, potențial datorită supraviețuirii crescute a celulelor nou diferențiate. Kougioumtzidou et al. (2017) au furnizat dovezi suplimentare că activitatea circuitului poate fi importantă în reglarea supraviețuirii celulelor – ei au demonstrat că pierderea subunităților 2, 3 și 4 ale receptorilor AMPA în OPC-uri duce la o supraviețuire redusă a oligodendrocitelor. Acest lucru sugerează că mielinizarea de novo ar putea fi modulată de activitatea circuitelor corticale pe tot parcursul vieții, poate pentru a regla fin funcția acelorași circuite.
Multe întrebări rămân de abordat: care este efectul oligodendrogenezei și al mielinizării de novo asupra funcției actuale a circuitului? Activitatea neuronală sporește supraviețuirea pe termen lung a oligodendrocitelor mielinizante? Este posibil ca pierderea oligodendrocitelor la bătrânețe să se datoreze reducerii activității neuronale asociate cu vârsta, ceea ce ar putea, la rândul său, să afecteze supraviețuirea generală a oligodendrocitelor. Alternativ, este posibil ca oligodendrocitele să aibă o durată de viață limitată, independent de activitatea neuronală (fie că sunt programate intrinsec, fie că sunt influențate de alte semnale extrinseci asociate cu îmbătrânirea). În oricare dintre cazuri, stimularea circuitului ar putea contribui la atenuarea pierderii de mielină asociată cu vârsta, fie prin promovarea supraviețuirii oligodendrocitelor existente, fie prin stimularea producției de noi oligodendrocite. Acest lucru, la rândul său, ar putea avea implicații semnificative în tratamentul și prevenirea declinului cognitiv asociat vârstei.
Oligodendrogeneza mediată de activitate nu este limitată la cortexul somatosenzorial – șoarecii adulți tineri supuși învățării motorii prezintă, de asemenea, o creștere a numărului de oligodendrocite nou diferențiate în cortexul motor (Xiao et al., 2016). Ce se întâmplă cu alte zone ale SNC? Mulți axoni corticali se proiectează prin corpul calos și, prin urmare, stimularea circuitelor corticale ar putea transmite semnale atât către OPC corticale, cât și către OPC calosale. Două studii pe rozătoare au demonstrat că stimularea neuronilor corticali induce oligodendrogeneza în corpul calos. Gibson et al. (2014) au stimulat optogenetic neuronii de proiecție din stratul V în cortexul premotor, constatând o creștere a proliferării OPC atât în cortexul premotor, cât și în corpul calos. Acest lucru a dus la o creștere a numărului de oligodendrocite și a grosimii tecii la 4 săptămâni după stimulare. Mai recent, Mitew et al. (2018) au folosit Receptorii de design activat exclusiv de medicamente de design pentru a stimula neuronii somatosenzoriali din stratul 2/3 și au observat, de asemenea, o creștere a proliferării OPC, oligodendrogeneză și teci de mielină mai groase în corpul calos atât la șoarecii tineri, cât și la cei adulți. Aceștia au demonstrat, de asemenea, că noile oligodendrocite formează în mod preferențial teci de mielină pe axonii activi. Acest lucru indică faptul că mielinizarea de novo indusă de activitate poate, în principiu, să vizeze axonii/circuitele active. Rămâne necunoscut cât de durabile ar putea fi modificările de durată ale mielinei ca răspuns la activitatea neuronală. Supraviețuirea pe termen lung a celulelor mielinizante observată de Tripathi et al. (2017), Hill et al. (2018) și Hughes et al. (2018) sugerează că, odată ce un oligodendrocit formează învelișuri de mielină, este probabil să supraviețuiască chiar dacă nivelurile de activitate neuronală revin înapoi la nivelul de bază. Dacă tecile de mielină în sine se schimbă odată ce activitatea neuronală revine la niveluri normale necesită mai multe investigații privind dinamica tecilor individuale, care este discutată mai jos.
Astfel, este posibil ca mielinizarea de novo pe tot parcursul vieții să apară în multe regiuni ale SNC, unde axonii potriviți pentru mielinizare au suficient spațiu nemielinizat. Cu toate acestea, rămâne neclar în ce măsură oligodendrogeneza continuă în diferite zone ale creierului uman adult. Analiza de datare cu carbon sugerează că majoritatea oligodendrocitelor din tractul corpului calos sunt generate în copilăria timpurie (Yeung et al., 2014). Analiza imunohistochimică a țesutului cerebral uman utilizând un nou marker pentru oligodendrocitele nou diferențiate (BCAS1) arată oligodendrocite noi în cortexul frontal chiar și după vârsta mijlocie, dar foarte puține oligodendrocite noi în substanța albă după al treilea deceniu de viață (Fard et al., 2017). Această diferență în oligodendrogeneza dintre specii ar putea fi un rezultat al scării. Datele obținute de Hughes et al. (2013) la rozătoare sugerează că oligodendrocitele sunt generate în exces uriaș, cu o tăiere continuă a aproape 80 % din celule. Având în vedere costul energetic al unui astfel de proces, este acest mecanism sustenabil pe tot parcursul vieții într-un organ de dimensiunea creierului uman? Poate că în creierul uman există o supraproducție limitată de oligodendrocite, din cauza necesității unei mielinizări mai îndelungate a SNC mai mare, sau pentru că semnalele, cum ar fi activitatea neuronală, stimulează OPC-urile să se diferențieze în oligodendrocite în funcție de necesități.
Remodelarea mielinei
Remodelarea tecilor de mielină existente ar putea modifica proprietățile de conducere fără a fi necesară generarea de noi oligodendrocite sau mielină. Modificarea lungimilor tecilor de mielină existente ar putea modifica acoperirea de mielină de-a lungul unui axon și distanța dintre nodurile Ranvier (care ar avea ambele un impact asupra vitezelor de conducere). În plus, chiar și remodelarea foarte subtilă a mielinei ar putea modifica lungimile nodurilor în sine. S-a demonstrat recent că lungimea nodurilor poate varia foarte mult în nervul optic și în cortex și că, în principiu, modificarea lungimii nodurilor de-a lungul unui axon poate, de asemenea, să modifice semnificativ viteza de conducere (Arancibia-Cárcamo et al., 2017). Rămâne de stabilit dacă modificările nodului Ranvier sunt determinate în primul rând de mielinizarea sau de reorganizarea axonului în sine.
Atât Hill et al. (2018), cât și Hughes et al. (2018) au efectuat un studiu longitudinal al tecilor de mielină individuale în cortexul somatosenzorial de șoarece timp de mai multe săptămâni pentru a evalua dacă lungimile tecilor sunt reglementate dinamic. Hill et al. (2018) au constatat că, la vârsta adultă timpurie (P90-120), deși unele teci prezintă extindere sau contracție, 81% dintre tecile observate au fost stabile. Este posibil ca mai multe teci să devină stabile în lungime odată cu vârsta; Hughes et al. (2018) au urmărit teci la animale mai în vârstă (P365) și au văzut că 99% dintre teci au rămas stabile pe parcursul a 3 săptămâni (Figura 1B).
O stabilitate similară a lungimii tecilor a fost, de asemenea, descrisă în altă parte; Auer et al. (2018) au folosit larve de pește zebră pentru a investiga dacă tecile individuale își pot schimba lungimea în timp, efectuând imagistica în direct în timp a tecilor de mielină marcate cu fluorescență. Ei au constatat că tecile individuale suferă o creștere rapidă, dar variabilă, în primele câteva zile după formare, înainte de a-și stabiliza lungimea tecilor. Odată stabilizate, tecile continuă să crească doar pentru a se adapta la creșterea generală a animalului.
De ce unele teci din cortex își schimbă lungimea, în timp ce altele nu? Acest lucru poate reflecta diversitatea în cerințele circuitelor neuronale distincte. Diversitatea axonală a fost observată în timpul mielinizării inițiale în măduva spinării peștelui zebră, unde unii axoni folosesc eliberarea veziculelor sinaptice pentru a regla numărul și lungimea tecii de mielină, în timp ce alții nu o fac (Koudelka et al., 2016). Acest lucru ridică ipoteza intrigantă că doar unii axoni sunt capabili să reglementeze mielina prin intermediul semnalelor legate de activitate. Hughes și colab. (2018) au constatat că paradigma lor de stimulare senzorială nu a crescut proporția de teci dinamice în cortexul somatosenzorial. Cu toate acestea, o analiză mai detaliată a diversității subtipurilor de axoni, împreună cu un studiu longitudinal al dinamicii lungimii tecii, ar putea confirma dacă remodelarea lungimii tecii este specifică anumitor circuite.
Stabilitatea lungimii tecii reflectă o incapacitate a tecilor de a se remodela? Experimentele la peștele zebră sugerează că remodelarea lungimii tecii poate fi indusă atunci când profilul de mielinizare al unui axon este perturbat. Auer et al. (2018) au ablaționat oligodendrocite individuale și, prin urmare, au îndepărtat în mod rarefiat tecile de-a lungul axonilor. Aceștia au constatat că, atunci când o singură teacă de mielină este pierdută pe un axon complet mielinizat, tecile vecine ar putea reincepe o creștere rapidă pentru a acoperi golul nemielinizat. În mai multe cazuri, o nouă teacă de mielină s-ar forma în locul predecesorului său și ar putea chiar împinge înapoi împotriva tecilor vecine invadatoare pentru a restabili modelul original de mielinizare (figura 2A). Prin urmare, uneori, un model specific de mielinizare este menținut în mod preferențial, chiar și după întreruperea mielinei. Acest lucru poate fi pentru a susține proprietățile de conducere optimizate ale axonului subiacent. Auer et al. (2018) au observat axoni slab mielinizați la larvele de pește zebră, așa cum a fost identificat anterior în cortexul rozătoarelor. În mod interesant, aceștia au constatat că, în urma ablației unor teci unice pe astfel de axoni slab mielinizați, o nouă teacă s-a format practic în același loc ca și teaca ablatată, chiar și de-a lungul unei porțiuni de axon altfel nemielinizate (Figura 2B). Astfel, modelele de mielinizare de-a lungul axonilor slab mielinizați par, de asemenea, să fie menținute în mod stabil la peștele zebră, așa cum au sugerat Hill et al. (2018) la rozătoare. Funcția profilurilor de mielinizare rarefiată rămâne necunoscută. Astfel de modele pot permite un reglaj fin mai dinamic al funcției unui singur axon în timp, deși este, de asemenea, posibil ca astfel de goluri nemielinizate să faciliteze mielinizarea treptată pentru a menține timpi de conducere consecvenți în cadrul circuitelor, pe măsură ce animalul crește și/sau lungimea axonilor se modifică.
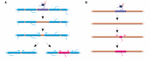
FIGURA 2. Remodelarea mielinei poate avea loc in vivo. (A) Ablația unor teci unice pe un axon complet mielinizat poate induce creșterea rapidă a tecilor învecinate pentru a acoperi golul. Acest decalaj poate fi acoperit în întregime de tecile învecinate sau profilul inițial de mielinizare poate fi restabilit prin adăugarea unei noi teci. (B) Ablația unui înveliș pe un axon slab mielinizat este urmată de formarea unui nou înveliș de mielină de dimensiuni și localizare identice cu învelișul predecesor ablatat. Rezumat al datelor din Auer et al. (2018).
Au și tecile stabile de mielină la mamifere această capacitate de remodelare atunci când modelul de mielinizare este perturbat? Pentru a răspunde la această întrebare sunt necesare studii longitudinale suplimentare cuplate cu demielinizarea. Este posibil ca o astfel de remodelare să nu fie indusă de activitatea neuronală, ci să fie un mecanism compensatoriu pentru pierderea de mielină. Pierderea oligodendrocitelor asociată vârstei ar putea declanșa remodelarea învelișurilor supraviețuitoare pentru a acoperi porțiunile denudate ale axonului și, prin urmare, să contribuie la menținerea funcției circuitului. Imagistica în direct a tecilor de mielină la bătrânețe ar putea determina dacă acesta este cazul.
Studiile de imagistică în direct discutate aici au evaluat toate dinamica lungimii tecii de mielină, dar nu și grosimea tecii de mielină. Poate fi reglementată dinamic grosimea tecii? Stimularea semnalizării PI3K/AKT/mTOR în oligodendrocitele șoarecilor adulți declanșează o înfășurare suplimentară a mielinei pentru a crește grosimea tecii (Snaidero et al., 2014). Acest lucru poate fi modulat de activitatea circuitului, deoarece stimularea neuronală duce la creșterea grosimii tecii atât la șoarecii tineri, cât și la cei adulți (Gibson et al., 2014; Mitew et al., 2018). Acest lucru evidențiază necesitatea de a obține imagini longitudinale ale tuturor parametrilor tecii pentru a înțelege pe deplin dinamica remodelării tecii. Este nevoie de modalități de imagistică în direct pentru a măsura cu exactitate grosimea tecii de-a lungul axonilor, deoarece, în prezent, acest lucru necesită măsurători transversale prin microscopie electronică, ceea ce limitează analiza la un singur punct temporal. Unele tehnici de imagistică fără etichete, cum ar fi microscopia de generare a treia armonică și reflectometria spectrală, se arată promițătoare pentru efectuarea unor astfel de măsurători (Lim et al., 2014; Kwon et al., 2017). Cuplarea acestor tehnici cu studii longitudinale ale cortexului rozătoarelor ar putea determina dacă tecile de mielină stabilite își pot ajusta grosimea sau dacă activitatea neuronală împinge pur și simplu mielinizarea de novo pentru a produce teci mai groase.
Se pare, prin urmare, că, deși tecile de mielină sunt capabile să se remodeleze atunci când mielina este întreruptă, majoritatea tecilor sunt în general stabile în lungime. Această stabilitate se datorează, potențial, menținerii modelelor de mielinizare stabilite timpuriu și optimizate pentru funcționarea circuitului.
Viitorul
Studiile recente de imagistică la mamifere s-au concentrat pe mielinizarea de novo și remodelarea tecilor în materia cenușie corticală. Circuitele corticale primesc și trimit informații prin intermediul mai multor regiuni, cum ar fi măduva spinării și corpul calos, astfel încât modificările mielinei în mai multe zone diferite ale SNC ar putea modifica semnalizarea într-un singur circuit. SNC este descris în mod tradițional în funcție de aspectul după fixarea în formaldehidă, unde „materia albă” descrie traiectele axonale puternic mielinizate, în timp ce „materia cenușie” descrie regiunile dens împachetate cu corpuri celulare neuronale, dendrite și sinapse. Cu toate acestea, această clasificare este prea simplistă; OPC-urile produc oligodendrocite mielinizante atât în materia cenușie, cât și în cea albă (Dawson et al., 2003) și, de fapt, există dovezi emergente de diversitate în linia oligodendroglială și în modelele de mielinizare atât în materia cenușie, cât și în cea albă (Rivers et al., 2008; Viganò et al., 2013; Young et al., 2013; Bechler et al., 2015). O astfel de diversitate poate reflecta cerințele unice ale mielinei în zone distincte și, potențial, pe circuite distincte ale SNC. Sunt necesare studii suplimentare de imagistică longitudinală pentru a înțelege mai bine dinamica mielinizării de novo și a remodelării învelișului în zone ale SNC dincolo de cortex.
În timp ce transparența optică a larvelor de pește zebră se pretează la imagistică neinvazivă în direct, efectuarea unor astfel de experimente în SNC de mamifere este mai invazivă și mai dificilă din punct de vedere tehnic. Hill et al. (2018) și Hughes et al. (2018) au utilizat microscopia cu doi fotoni cu ferestre de imagistică craniană pentru a obține imagini la adâncimi de până la 400 μm în cortex. Tehnici similare ar putea fi utilizate pentru a imagina în timp traiectele mielinizate superficiale din măduva spinării (Locatelli et al., 2018), dar regiunile mai profunde ale SNC nu pot fi penetrate doar prin microscopie cu doi fotoni. O alternativă este utilizarea microendoscopiei cu doi fotoni, în care o sondă microendoscopică cu o lentilă cu indice de refracție gradient (GRIN) este introdusă în țesut pentru a imagina celulele mai adânc în creier . Cu toate acestea, inserția endoscopului poate duce la răspunsuri inflamatorii care ar putea avea un impact asupra mielinizării. O alternativă ar putea fi microscopia cu trei fotoni utilizând metoda ferestrei de imagistică craniană, care a fost, de asemenea, utilizată anterior pentru a obține imagini ale hipocampusului (Horton et al., 2013; Ouzounov et al., 2017). Microscopia cu trei fotoni oferă un raport semnal-fond semnificativ mai mare decât microscopia cu doi fotoni și, prin urmare, poate fi utilizată pentru a imagina structuri tisulare mai profunde.
Este deosebit de important să se ia în considerare nu numai diferite regiuni ale SNC, ci și diferiți neuroni din aceste regiuni. Cercetările anterioare sugerează că există diferențe mecaniciste în modul în care subtipurile distincte de neuroni își reglează mielinizarea (Koudelka et al., 2016). În plus, este posibil să existe diversitate în reglarea locală a mielinei. Este esențial să ne amintim că diferitele părți ale SNC nu sunt entități separate, ci sunt interconectate. Integrarea connectomicii la mezoscale, care se concentrează pe înțelegerea conexiunilor diferitelor subtipuri de neuroni din diferite regiuni (Zeng, 2018), va fi crucială pentru înțelegerea modului în care dinamica mielinizării pe tot parcursul vieții variază între diferite circuite.
Care este consecința funcțională a reglării mielinei de-a lungul circuitelor distincte? Până în prezent, implicațiile funcționale pot fi deduse doar prin corelații cu comportamentul. În cele din urmă, este necesar să se cupleze măsurarea dinamicii mielinei cu evaluarea directă a activității circuitelor. Acest lucru va necesita înregistrarea activității neuronale în timpul studiilor longitudinale de mielinizare, pentru a conecta direct mielinizarea de novo sau remodelarea tecii observate cu modificările în funcție de circuit în timp. Va fi important să se măsoare dinamica mielinei și activitatea electrofiziologică a neuronilor și axonilor individuali pentru a determina modul în care modificările diferiților parametri ai învelișului de mielină afectează de fapt proprietățile de conducere la nivelul unei singure celule, precum și pentru a evalua activitatea la nivelul unei populații. Instrumente precum indicatorii de Ca2+ sau de tensiune codificați genetic permit înregistrarea relativ neinvazivă a activității circuitelor și pot fi utilizați chiar pentru a evalua activitatea circuitelor la nivelul întregului creier (Ahrens et al., 2012; Lovett-Barron et al., 2017).
Concluzie
Mielinizarea axonilor reprezintă un mecanism potențial puternic de reglare a funcției circuitelor pe tot parcursul vieții. Cercetările au demonstrat că mielinizarea de novo în cortex (prin producerea de noi oligodendrocite) are loc chiar și la vârsta adultă și că aceasta poate fi îmbunătățită prin stimularea activității circuitului. Odată ce s-a format mielina, aceasta este stabilă, cu o reînnoire redusă a oligodendrocitelor și o remodelare limitată a lungimilor tecilor de mielină existente. Cu toate acestea, aceste structuri stabile pot păstra capacitatea de remodelare în cazul în care mielina este perturbată. Acest lucru are implicații interesante în ceea ce privește plasticitatea mielinei în menținerea funcției circuitelor în timpul leziunilor, bolilor și bătrâneții. Rămâne de văzut cu exactitate modul în care modificările în mielinizare afectează funcția circuitului de bază. În cele din urmă, este necesară o abordare la nivel de circuit, care să integreze analiza dinamicii mielinei cu măsurarea directă a funcției circuitului, pentru a aprecia pe deplin modul în care mielinizarea dinamică influențează funcția generală a sistemului nervos pe tot parcursul vieții.
Contribuții ale autorilor
Toți autorii enumerați au avut o contribuție substanțială, directă și intelectuală la această lucrare și au aprobat-o pentru publicare.
Finanțare
DAL este susținută de o bursă de cercetare Wellcome Trust Senior Research Fellowship (102836/Z/13/Z). JMW este susținut de o bursă de doctorat la Universitatea din Edinburgh (MRC Doctoral Training Partnership MR/K501293/1) și de Wellcome Trust Four-Year Ph.D. Program in Tissue Repair (Grant 108906/Z/15/Z).
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Ahrens, M. B., Li, J. M., Orger, M. B., Robson, D. N., Schier, A. F., Engert, F., et al. (2012). Dinamica neuronală la nivelul creierului în timpul adaptării motorii la peștele zebră. Nature 485, 471-477. doi: 10.1038/nature11057
PubMed Abstract | CrossRef Full Text | Google Scholar
Almeida, R. G., și Lyons, D. A. (2017). Despre plasticitatea axonului mielinizat și formarea și funcția circuitului neuronal. J. Neurosci. 37, 10023-10034. doi: 10.1523/JNEUROSCI.3185-16.2017
CrossRef Full Text | Google Scholar
Arancibia-Cárcamo, I. L., Ford, M. C., Cossell, L., Ishida, K., Tohyama, K., și Attwell, D. (2017). Lungimea nodului Ranvier ca un potențial regulator al vitezei de conducere a axonului mielinizat. eLife 6:e23329. doi: 10.7554/eLife.23329
PubMed Abstract | CrossRef Full Text | Google Scholar
Auer, F., Vagionitis, S., și Czopka, T. (2018). Dovezi pentru remodelarea tecii de mielină în SNC relevate de imagistica in vivo. Curr. Biol. 28, 549-559. doi: 10.1016/j.cub.2018.01.017
PubMed Abstract | CrossRef Full Text | Google Scholar
Barres, B. A., Hart, I. K., Coles, H. S. R., Burne, J. F., Voyvodic, J. T., Richardson, W. D., et al. (1992). Moartea celulară și controlul supraviețuirii celulelor în linia oligodendrocitelor. Cell 70, 31-46. doi: 10.1016/0092-8674(92)90531-G
CrossRef Full Text | Google Scholar
Bechler, M. E., Byrne, L., and Ffrench-Constant, C. (2015). Lungimile tecii de mielină din SNC sunt o proprietate intrinsecă a oligodendrocitelor. Curr. Biol. 25, 2411-2416. doi: 10.1016/j.cub.2015.07.056
PubMed Abstract | CrossRef Full Text | Google Scholar
Chang, A., Nishiyama, A., Peterson, J., Prineas, J., și Trapp, B. D. (2000). NG2-pozitive NG2-pozitive oligodendrocyte progenitor cells in adult human brain and multiple sclerosis lesions. J. Neurosci. 20, 6404-6412. doi: 10.1523/JNEUROSCI.20-17-06404.2000
PubMed Abstract | CrossRef Full Text | Google Scholar
Cox, S. R., Ritchie, S. J., Ritchie, S. J., Tucker-Drob, E. M., Liewald, D. C., Hagenaars, S. P., Davies, G., et al. (2016). Îmbătrânirea și structura materiei albe a creierului la 3.513 participanți la UK Biobank. Nat. Commun. 7:13629. doi: 10.1038/ncomms13629
PubMed Abstract | Reflect Full Text | Google Scholar
Czopka, T., Ffrench-Constant, C., și Lyons, D. A. (2013). Oligodendrocitele individuale au doar câteva ore în care pot genera noi teci de mielină in vivo. Dev. Cell 25, 599-609. doi: 10.1016/j.devcel.2013.05.013
PubMed Abstract | CrossRef Full Text | Google Scholar
Dawson, M. R. L., Polito, A., Levine, J. M., și Reynolds, R. (2003). NG2-exprimând celulele progenitoare gliale NG2: o populație abundentă și răspândită de celule ciclice în CNS de șobolan adult. Mol. Cell. Neurosci. 24, 476-488. doi: 10.1016/S1044-7431(03)00210-0
PubMed Abstract | Refef Full Text | Google Scholar
Fard, M. K., Van der Meer, F., Sánchez, P., Cantuti-Castelvetri, L., Mandad, S., Jäkel, S., et al. (2017). Expresia BCAS1 definește o populație de oligodendrocite mielinizante timpurii în leziunile de scleroză multiplă. Sci. Transl. Med. 9:eaam7816. doi: 10.1126/scitranslmed.aam7816
PubMed Abstract | CrossRef Full Text | Google Scholar
Fields, R. D. (2015). Un nou mecanism de plasticitate a sistemului nervos: mielinizarea dependentă de activitate. Nat. Rev. Neurosci. 16, 756-767. doi: 10.1038/nrn4023
PubMed Abstract | Refef Full Text | Google Scholar
Ford, M. C., Alexandrova, O., Cossell, L., Stange-Marten, A., Sinclair, J., Kopp-Scheinpflug, C., et al. (2015). Reglarea proprietăților nodului Ranvier și a internodurilor în axonii mielinizați pentru a ajusta calendarul potențialului de acțiune. Nat. Commun. 6:8073. doi: 10.1038/ncomms9073
PubMed Abstract |Ref Full Text | Google Scholar
Gibson, E. M., Purger, D., Mount, C. W., Goldstein, A. K., Lin, G. L., Wood, L. S., et al. (2014). Activitatea neuronală promovează oligodendrogeneza și mielinizarea adaptivă în creierul mamiferelor. Science 344:1252304. doi: 10.1126/science.1252304
PubMed Abstract | Text integral | Google Scholar
Hill, R. A., Li, A. M., și Grutzendler, J. (2018). Plasticitatea mielinei corticale pe tot parcursul vieții și degenerarea legată de vârstă în creierul viu de mamifere. Nat. Neurosci. 21, 683-695. doi: 10.1038/s41593-018-0120-6
PubMed Abstract | Refef Full Text | Google Scholar
Hill, R. A., Patel, K. D., Goncalves, C. M., Grutzendler, J., și Nishiyama, A. (2014). Modularea generării oligodendrocitelor în timpul unei ferestre temporale critice după diviziunea celulelor NG2. Nat. Neurosci. 17, 1518-1527. doi: 10.1038/nn.3815
PubMed Abstract | Text integral | Google Scholar
Horton, N. G., Wang, K., Kobat, D., Clark, C. G., Wise, F. W., Schaffer, C. B., et al. (2013). Microscopia cu trei fotoni in vivo a structurilor subcorticale în cadrul unui creier de șoarece intact. Nat. Photonics 7, 205-209. doi: 10.1038/NPHOTON.2012.336
PubMed Abstract | Full CrossRef Text | Google Scholar
Huang, B., Wei, W., Wang, G., Gaertig, M. A., Feng, Y., Wang, W., et al. (2015). Huntingtinul mutant reglează în jos expresia genei de mielină mediată de factorul de reglementare a mielinei și afectează oligodendrocitele mature. Neuron 85, 1212-1226. doi: 10.1016/j.neuron.2015.02.026
PubMed Abstract | CrossRef Full Text | Google Scholar
Hughes, E. G., Kang, S. H., Fukaya, M., și Bergles, D. E. (2013). Progenitorii de oligodendrocite echilibrează creșterea cu auto-repulsia pentru a realiza homeostazia în creierul adult. Nat. Neurosci. 16, 668-676. doi: 10.1038/nn.3390
PubMed Abstract | Textul integral | Google Scholar
Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J. J., și Bergles, D. E. (2018). Remodelarea mielinei prin oligodendrogeneza dependentă de experiență în cortexul somatosenzorial adult. Nat. Neurosci. 21, 696-706. doi: 10.1038/s41593-018-0121-5
PubMed Abstract | Full CrossRef Text | Google Scholar
Jung, J. C., Mehta, A. D., Aksay, E., Stepnoski, R., și Schnitzer, M. J. (2004). Imagistica in vivo a creierului mamiferelor folosind microendoscopia cu fluorescență cu unul și doi fotoni. J. Neurophysiol. 92, 3121-3133. doi: 10.1152/jn.00234.2004
PubMed Abstract | CrossRef Full Text | Google Scholar
Kang, S. H., Li, Y., Fukaya, M., Lorenzini, I., Cleveland, D. W., Ostrow, L. W., et al. (2013). Degenerarea și regenerarea afectată a oligodendrocitelor din materia cenușie în scleroza laterală amiotrofică. Nat. Neurosci. 16, 571-579. doi: 10.1038/nn.3357
PubMed Abstract |Ref Full Text | Google Scholar
Koudelka, S., Voas, M. G., Almeida, R. G., Baraban, M., Soetaert, J., Meyer, M. P., et al. (2016). Subtipurile neuronale individuale prezintă diversitate în mielinizarea SNC mediată de eliberarea veziculelor sinaptice. Curr. Biol. 26, 1447-1455. doi: 10.1016/j.cub.2016.03.070
PubMed Abstract | Ref Full Text | Google Scholar
Kougioumtzidou, E., Shimizu, T., Hamilton, N. B., Tohyama, K., Sprengel, R., Monyer, H., et al. (2017). Semnalizarea prin intermediul receptorilor AMPA pe precursorii oligodendrocitelor promovează mielinizarea prin creșterea supraviețuirii oligodendrocitelor. eLife 6:e2808080. doi: 10.7554/eLife.28080
PubMed Abstract | PublicMed Abstract | Google Scholar
Kwon, J., Kim, M., Park, H., Kang, B.-M., Jo, Y., Kim, J.-H., et al. (2017). Metrologie optică la scară nanometrică fără etichetă pe axoni mielinizați in vivo. Nat. Commun. 8:1832. doi: 10.1038/s41467-017-01979-2
PubMed Abstract | Text integral | Google Scholar
Levene, M. J., Dombeck, D. A., Kasischke, K. A., Molloy, R. P., și Webb, W. W. (2004). In vivo multiphoton microscopy of deep brain tissue. J. Neurophysiol. 91, 1908-1912. doi: 10.1152/jn.01007.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Lim, H., Sharoukhov, D., Kassim, I., Zhang, Y., Salzer, J. L., și Melendez-Vasquez, C. V. (2014). Imagistica fără etichetă a mielinizării celulelor Schwann prin microscopie de generație armonică a treia. Proc. Natl. acad. Sci. U.S.A. 111, 18025-18030. doi: 10.1073/pnas.1417820111
PubMed Abstract | CrossRef Full Text | Google Scholar
Locatelli, G., Theodorou, D., Kendirli, A., Jordão, M. J. C., Staszewski, O., Phulphagar, K., et al. (2018). Fagocitele mononucleare își specifică și își adaptează fenotipul la nivel local într-un model de scleroză multiplă. Nat. Neurosci. 21, 1196-1208. doi: 10.1038/s41593-018-0212-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovett-Barron, M., Andalman, A. S., Allen, W. E., Vesuna, S., Kauvar, I., Burns, V. M., et al. (2017). Circuite ancestrale pentru modularea coordonată a stării creierului. Cell 171, 1411-1423. doi: 10.1016/j.cell.2017.10.021
PubMed Abstract | Text integral | Google Scholar
Mabbott, D. J., Noseworthy, M., Bouffet, E., Laughlin, S., și Rockel, C. (2006). Creșterea materiei albe ca mecanism de dezvoltare cognitivă la copii. Neuroimage 33, 936-946. doi: 10.1016/j.neuroimage.2006.07.024
PubMed Abstract | CrossRef Full Text | Google Scholar
McKenzie, I. A., Ohayon, D., Li, H., Paes de Faria, J., Emery, B., Tohyama, K., et al. (2014). Învățarea abilităților motorii necesită mielinizare centrală activă. Science 346, 318-322. doi: 10.1126/science.1254960
PubMed Abstract | Full CrossRef Text | Google Scholar
Mitew, S., Gobius, I., Fenlon, L. R., McDougall, S. J., Hawkes, D., Xing, Y. L., et al. (2018). Stimularea farmacogenetică a activității neuronale crește mielinizarea într-o manieră specifică axonului. Nat. Commun. 9:306. doi: 10.1038/s41467-017-02719-2
PubMed Abstract | Full CrossRef Text | Google Scholar
Mount, C. W., și Monje, M. (2017). Înfășurat pentru a se adapta: mielinizarea dependentă de experiență. Neuron 95, 743-756. doi: 10.1016/j.neuron.2017.07.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Ouzounov, D. G., Wang, T., Wang, M., Feng, D. D. D., Horton, N. G., Cruz-Hernández, J. C., et al. (2017). Imagistica in vivo cu trei fotoni a activității neuronilor marcați cu GCaMP6 în profunzime în creierul intact de șoarece. Nat. Methods 14, 388-390. doi: 10.1038/nmeth.4183
PubMed Abstract | Refef Full Text | Google Scholar
Ritchie, S. J., Bastin, M. E., Tucker-Drob, E. M. M., Maniega, S. M., Engelhardt, L. E., Cox, S. R., et al. (2015). Modificări cuplate în microstructura materiei albe a creierului și inteligența fluidă la vârsta a doua. J. Neurosci. 35, 8672-8682. doi: 10.1523/JNEUROSCI.0862-15.2015
PubMed Abstract | Cross Full Text | Google Scholar
Rivers, L. E., Young, K. M., Rizzi, M., Jamen, F., Psachoulia, K., Wade, A., et al. (2008). Glia PDGFRA/NG2 generează oligodendrocite mielinizante și neuroni de proiecție piriformă la șoarecii adulți. Nat. Neurosci. 11, 1392-1401. doi: 10.1038/nn.2220
PubMed Abstract | Textul integral | Google Scholar
Sampaio-Baptista, C., Khrapitchev, A. A., Foxley, S., Schlagheck, T., Scholz, J., Jbabdi, S., et al. (2013). Învățarea abilităților motorii induce modificări în microstructura materiei albe și mielinizarea. J. Neurosci. 33, 19499-19503. doi: 10.1523/jneurosci.3048-13.2013
PubMed Abstract | CrossRef Full Text | Google Scholar
Scantlebury, N., Cunningham, T., Dockstader, C., Laughlin, S., Gaetz, W., Rockel, C., et al. (2014). Relațiile dintre maturizarea materiei albe și timpul de reacție în copilărie. J. Int. Neurophsychol. Soc. 20, 99-112. doi: 10.1017/S1355617713001148
PubMed Abstract | CrossRef Full Text | Google Scholar
Schain, A. J., Hill, R. A. A., și Grutzendler, J. (2014). Imagistica in vivo fără etichetă a axonilor mielinizați în sănătate și boală cu ajutorul microscopiei confocale spectrale de reflectanță. Nat. Med. 20, 443-449. doi: 10.1038/nm.3495
PubMed Abstract |Ref Full Text | Google Scholar
Scholz, J., Klein, M. C., Behrens, T. E. J., și Johansen-berg, H. (2009). Antrenamentul induce schimbări în arhitectura materiei albe. Nat. Neurosci. 12, 1370-1371. doi: 10.1038/nn.2412
PubMed Abstract | Textul integral | Google Scholar
Snaidero, N., Möbius, W., Czopka, T., Hekking, L. H. P., Mathisen, C., Verkleij, D., et al. (2014). Înfășurarea membranei de mielină a axonilor CNS prin creșterea polarizată dependentă de PI(3,4,5)P3 la nivelul limbii interne. Cell 156, 277-290. doi: 10.1016/j.cell.2013.11.044
PubMed Abstract | Text integral | Google Scholar
Takahashi, N., Sakurai, T., Davis, K. L., și Buxbaum, J. D. (2011). Legarea disfuncției oligodendrocitelor și a mielinei la anomaliile neurocircuitelor în schizofrenie. Prog. Neurobiol. 93, 13-24. doi: 10.1016/j.pneurobio.2010.09.004
PubMed Abstract | Full CrossRef Text | Google Scholar
Tomassy, G. S., Berger, D. R., Chen, H.-H., Kasthuri, N., Hayworth, K. J., Vercelli, A., et al. (2014). Profiluri distincte de distribuție a mielinei de-a lungul axonilor individuali ai neuronilor piramidali în neocortex. Science 344, 319-324. doi: 10.1126/science.1249766
PubMed Abstract | Text integral | Google Scholar
Tripathi, R. B., Jackiewicz, M., Mckenzie, I. A., Kougioumtzidou, E., Grist, M., și Richardson, W. D. (2017). Stabilitatea remarcabilă a oligodendrocitelor mielinizante la șoareci. Cell Rep. 21, 316-323. doi: 10.1016/j.celrep.2017.09.050
PubMed Abstract | CrossRef Full Text | Google Scholar
Viganò, F., Möbius, W., Götz, M., și Dimou, L. (2013). Transplantul dezvăluie diferențe regionale în diferențierea oligodendrocitelor în creierul adult. Nat. Neurosci. 16, 1370-1372. doi: 10.1038/nn.3503
PubMed Abstract | Textul integral | Google Scholar
Watkins, T. A., Emery, B., Mulinyawe, S., și Barres, B. A. (2008). Etape distincte de mielinizare reglementate de gamma-secretase și astrocite într-un sistem de cocultură a SNC cu mielinizare rapidă. Neuron 60, 555-569. doi: 10.1016/j.neuron.2008.09.011
PubMed Abstract | CrossRef Full Text | Google Scholar
Waxman, S. G. (1980). Determinanți ai vitezei de conducere în fibrele nervoase mielinizate. Muscle Nerve 3, 141-150. doi: 10.1002/mus.880030207
PubMed Abstract | Full Text | Google Scholar
Xiao, L., Ohayon, D., Mckenzie, I. A., Sinclair-Wilson, A., Wright, J. L., Fudge, A. D., et al. (2016). Producția rapidă de noi oligodendrocite este necesară în cele mai timpurii etape ale învățării abilităților motorii. Nat. Neurosci. 19, 1210-1217. doi: 10.1038/nn.4351
PubMed Abstract | Text integral | Google Scholar
Yeung, M. S. Y., Zdunek, S., Bergmann, O., Bernard, S., Salehpour, M., Alkass, K., et al. (2014). Dinamica generării oligodendrocitelor și a mielinizării în creierul uman. Cell 159, 766-774. doi: 10.1016/j.cell.2014.10.011
PubMed Abstract | Full CrossRef Text | Google Scholar
Young, K. M., Psachoulia, K., Tripathi, R. B., Dunn, S.-J., Cossell, L., Attwell, D., et al. (2013). Dinamica oligodendrocitelor în SNC adult sănătos: dovezi pentru remodelarea mielinei. Neuron 77, 873-885. doi: 10.1016/j.neuron.2013.01.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Zeng, H. (2018). Conexomica la mezoscală. Curr. Opin. Neurobiol. 50, 154-162. doi: 10.1016/j.conb.2018.03.003
PubMed Abstract | Full Text | Google Scholar
.